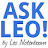This textbook will cover ways to communicate effectively as you develop insight into your own style, writing process,
this book will be helpful to future refrences
This textbook will cover ways to communicate effectively as you develop insight into your own style, writing process,
this book will be helpful to future refrences
CNM students have access to The Learning and Computer Center (TLCc), which is available on six campuses: Advanced Technology Center, Main, Montoya, Rio Rancho, South Valley, and Westside. At these writing centers, trained tutors help students meet college-level expectations. The tutoring centers offer one-on-one meetings, online, and group sessions for multiple disciplines. TLCc also offers workshops on citing and learning how to develop a writing process.
Lists campus locations. Offers: One-on-one tutoring Online sessions Workshops Supports multiple disciplines.
Communication courses teach students that communication involves two parties—the sender and the receiver of the communicated message. Sometimes, there is more than one sender and often, there is more than one receiver of the message. The main purpose of communication whether it be email, text, tweet, blog, discussion, presentation, written assignment, or speech is always to help the receiver(s) of the message understand the idea that the sender of the message is trying to share. This section will focus on electronic communication in a college course.
This textbook was designed for English 1110 and 1120, Composition I and Composition II, respectively. If you are enrolled in one of these courses, you may be nearing the end of your studies at Central New Mexico Community College (CNM)
This textbook is specifically created for CNM Composition I & II students
Don’t Trust the Cloud. You Need to Back Up Your Email and Online DataTap to unmute2xDon’t Trust the Cloud. You Need to Back Up Your Email and Online DataAsk Leo! 12,913 views 10 months agoSearchCopy linkInfoShoppingIf playback doesn't begin shortly, try restarting your device.Pull up for precise seekingMute2:18How vulnerable are you?•You're signed outVideos that you watch may be added to the TV's watch history and influence TV recommendations. To avoid this, cancel and sign in to YouTube on your computer.CancelConfirmIf you get an answer at all, that is.Up nextLiveUpcomingCancelPlay nowAsk Leo!SubscribeSubscribedVideos from Ask Leo! - https://askleo.com Answers in plain language, presented so you can understand. I help you use technology more effectively, giving you the confidence to do more, be more, and explore more of the world that today's technology makes available to us all. Making Technology Work For Everyone - Ask Leo!askleo.comVisitIs It Safe to Look, Just Look, at Spam?6:16HideShareInclude playlistAn error occurred while retrieving sharing information. Please try again later.0:011:31 / 9:09Live•Watch full video•Online providers don’t have your back•8:31A Father’s Last Goodbye? Michael Bennett Performs “I Just Might” on America’s Got TalentAGTverse11k views • 8 hours agoLivePlaylist ()Mix (50+)20:02Voice Coach/Opera Singer REACTION & ANALYSIS Disturbed "The Sound of Silence"The Charismatic Voice8.3m views • 5 years agoLivePlaylist ()Mix (50+)4:16LESSON 1: IP ADDRESSES ≠ IDENTITY (THE BASICS)#VideoJesus183 views • 2 weeks agoLivePlaylist ()Mix (50+)12:03I’ve Stopped Using Google Search! Why you should consider it too.Ask Leo!1m views • 1 year agoLivePlaylist ()Mix (50+)9:32Why Orban’s Fall Looks InevitableTLDR News EU256k views • 1 day agoLivePlaylist ()Mix (50+)17:44Feynman: Why The Universe Obeys "Invented" MathThe Feynman Lens53k views • 5 days agoLivePlaylist ()Mix (50+)15:30I Fried ChatGPT With ONE Simple QuestionRick Beato976k views • 4 months agoLivePlaylist ()Mix (50+)15:54How Do I Store Files on My Computer and Not OneDrive?Ask Leo!566k views • 11 months agoLivePlaylist ()Mix (50+)4:51Michael Bennett on America’s Got Talent sings Cold Hands, Warm Heart and breaks hearts live onstage!Tears and Talents250k views • 3 days agoLivePlaylist ()Mix (50+)7:06The Fall of Bill GatesNewsthink464k views • 13 days agoLivePlaylist ()Mix (50+)29:09Bruce Lee Was Mocked by a Navy SEAL Who Said "Come Fight a Real Man" Only 8 Witnessed ItBruce Lee Encounters902k views • 3 weeks agoLivePlaylist ()Mix (50+)29:14Secretary Rubio delivers remarks to the Munich Security ConferenceU.S. Department of State610k views • 6 days agoLivePlaylist ()Mix (50+) Don’t Trust the Cloud. You Need to Back Up Your Email and Online Data
💻.asus.🧊.♖-05xy
IPNS Publishing keys are given personal names referred to as inter planetary personal names ippns
these names are made up by the creator of the file system and show the path to the folder that would be the root of the mutable filesystem that is being named followed by a dash and the last four character taken from the public key
💻.asus.🧊.♖-05xy
the personally understood name goves tha name of the machine used as the root elements in the file system 💻.asus. the ice icon signifies that this folderis part of an IPFS file system
the white rook utf8 icon ♖signifies that a peergos file system is being mirrored here in 🧊 IPFS
So retrieval depends on the storage infrastructure technology-protocol chosen But these can be global unique names, and there may be others but the combination of these two is self-sufficient
IPNS Publishing Keys
IPNS Publishing Key Names compreise two parts
the first igives the path to the folder that is to be published such that the / are replaced by dots . followed by a dash and the last four or five characters from the end of the public key
Xie, Min, Markus Eyting, Christian Bommer, Haroon Ahmed, and Pascal Geldsetzer. 2025. “The Effect of Shingles Vaccination at Different Stages of the Dementia Disease Course.” Cell 188(25): 7049-7064.e20. https://www.cell.com/cell/fulltext/S0092-8674(25)01256-5 (February 20, 2026).
Shallow understanding from people of good will is more frustrating than absolute misunderstanding from people of ill will. Lukewarm acceptance is much more bewildering than outright rejection.
I am able to record its considerable wisdom, shown in evading the plots of its enemy the spider. It is always on the look-out for his ambushes, and in the most circumspect way dodges about, that it may not be caught, netted, and entangled in his meshes
Agarra cualquier parecido con el humano, primero a nivel fisioólogico y después expande a elementos y cultura compartida con con alos seres vivos.
En particular la dinámica de depresdadores y presas.
Portrait de la santé mentale chez les jeunes : Analyse et perspectives du Mouvement Jeunes et santé mentale
Ce document synthétise les conclusions du « Portrait » mené par le Mouvement Jeunes et santé mentale (MJSM), une initiative de participation citoyenne impliquant plus de 850 jeunes Québécois âgés de 14 à 35 ans.
L'analyse révèle une fracture profonde entre les besoins réels des jeunes et les réponses institutionnelles actuelles, marquées par une médicalisation excessive et une approche fragmentée des soins.
Les points saillants incluent :
• Une demande de normalisation : Les jeunes revendiquent le droit de vivre et d'exprimer leur détresse sans être systématiquement pathologisés ou stigmatisés.
• La réalité numérique : Pour cette génération, l'espace virtuel n'est pas distinct de la « vraie vie », mais constitue un continuum essentiel pour le soutien et l'identité.
• L'échec de l'accès aux soins : Le système actuel impose des barrières géographiques, linguistiques et diagnostiques qui découragent la recherche d'aide.
• L'urgence d'un filet social : Face à une perte d'espoir collectif, le mouvement lance la campagne « Fini de patcher, temps de tisser serré », exigeant des solutions collectives plutôt que des interventions individuelles superficielles.
--------------------------------------------------------------------------------
Le Mouvement Jeunes et santé mentale, récemment incorporé, est né en 2016 d'une initiative de trois regroupements fondateurs (RRASMQ, Auberges du cœur, AGDSMQ), rejoints plus tard par le ROCLD et le ROCAJ.
Sa mission est de lutter collectivement contre la médicalisation des difficultés vécues par les jeunes.
Le MJSM se distingue par une gouvernance où les jeunes sont les décideurs et l'équipe permanente l'exécutant.
Le Portrait a été coconstruit selon trois valeurs fondamentales :
1. Approche sensible : Écoute active sans jugement, évitant de retraumatiser par des questions intrusives.
2. Intersectionnalité : Prise en compte de la pluralité des oppressions (classe, race, genre, handicap).
3. Processus collectif : Priorité donnée à la parole des jeunes à chaque étape.
| Outils de collecte | Données recueillies | | --- | --- | | Questionnaires en ligne | Plus de 440 réponses (dont 90 % complètes malgré le caractère facultatif). | | Ateliers en ressources jeunesse | Réalisés dans 12 régions du Québec. | | Focus groups ciblés | Jeunes issus de la DPJ, racisés, neurodivergents, LGBTQ+, en situation de handicap physique. | | Total des jeunes consultés | Plus de 850 individus. |
--------------------------------------------------------------------------------
Le Portrait est structuré en huit cahiers thématiques abordant les enjeux cruciaux identifiés par les participants.
Les jeunes rejettent la vision binaire de la santé mentale. Ils affirment que la détresse est une réaction normale au contexte actuel.
• Revendication : Normaliser sans banaliser.
Il s'agit d'accepter l'expression d'émotions dites négatives (colère, tristesse) sans vouloir les « régler » immédiatement par une pilule ou un diagnostic.
• Besoin d'écoute : Un désir massif de pouvoir s'exprimer dans des espaces sécurisés (écoles, familles) sans conséquences négatives ou jugements.
Les jeunes font une différence nette entre deux types de réseaux :
• Groupe d'appartenance : Des pairs qui partagent les mêmes réalités (souvent en ligne), essentiels pour la validation émotionnelle.
• Réseau de soutien concret : Les personnes capables d'aider lors d'une crise matérielle (prêter de l'argent, aider à un déménagement).
Le manque de soutien concret transforme souvent une détresse passagère en crise majeure.
Contrairement aux générations précédentes, les jeunes ne séparent pas le virtuel du réel.
• Avantages : Accès à l'information, représentation des identités marginalisées, premier contact moins menaçant avec les ressources.
• Risques : Chambres d'écho, haine en ligne, comparaison sociale.
• Recommandation : Les jeunes souhaitent que les professionnels intègrent le numérique comme un outil de transition vers les services en personne, plutôt que de s'y opposer.
Le titre, suggéré par les jeunes, souligne que le système actuel crée activement des barrières.
• Critique du diagnostic : L'obligation d'avoir un diagnostic pour accéder aux soins est perçue comme un frein majeur.
• Érance médicale : Les jeunes sont fatigués de répéter leur histoire en silo à des professionnels qui ne communiquent pas entre eux.
• Rapport de force : Une crainte persistante de se voir imposer des solutions sans être consulté.
--------------------------------------------------------------------------------
La consultation des plus jeunes a mis en lumière des anxiétés précoces liées à la performance et à l'avenir.
• Milieu scolaire : Pression étouffante des notes et sentiment de détachement de la part des professionnels (profs et psychologues) perçus comme débordés.
• Anxiété face à l'avenir : Des jeunes de 13 ans s'inquiètent déjà de la fiscalité, du logement et de leur capacité à trouver un emploi.
• Espaces de décompression : Un besoin criant de « safe spaces » à l'école où il est permis d'exister sans objectif de performance.
--------------------------------------------------------------------------------
L'analyse des données a forcé le MJSM à élargir ses revendications initiales, trop centrées sur les services techniques.
« Le droit de vivre des périodes de détresse doit être pleinement reconnu, sans culpabilisation ni conséquences négatives. »
Le mouvement constate une perte d'espoir collectif : les jeunes ne croient plus que les décideurs prennent en compte leur avenir.
Campagne : « Fini de patcher, temps de tisser serré »
Cette campagne vise à transformer l'indignation en force politique pour dénoncer la destruction du filet social.
| Objectif | Action prévue | | --- | --- | | Responsabilisation | Pressions auprès des élus pour inclure le financement de solutions collectives dans les plateformes électorales. | | Visibilité | Utilisation d'un symbole commun (un « plaster » en feu) pour solidariser les luttes. | | Mobilisation | Lancement public le 16 avril avec une trousse d'outils pour les groupes communautaires. | | Alternative | Promouvoir des approches basées sur la transformation sociale plutôt que sur la gestion individuelle de l'anxiété. |
• Sur la médicalisation : « On n’avait pas besoin de pilules, on avait besoin d’aide, on avait besoin qu’on m’écoute. »
• Sur le filet social : « L'absurdité de militer pour des miettes pendant que la maison brûle. »
• Sur la confidentialité : Les jeunes ne veulent pas d'un secret absolu, mais du « contrôle sur le partage de leur information » pour éviter de répéter sans cesse leur trauma.
En conclusion, le Mouvement Jeunes et santé mentale appelle à un changement de paradigme : passer d'une gestion de crise individuelle à un investissement massif dans des solutions collectives inclusives et porteuses d'espoir.
This Is How a Child Dies of Measles<br /> by [[Elizabeth Bruenig]] in The Atlantic<br /> accessed on 2026-02-20T09:59:04
keep unvaccinated children at home for the next three weeks, the virus’s maximum incubation period.
Among the unvaccinated, one in five people infected with measles in the United States will require hospitalization, and roughly two out of every 1,000 infected children will die of complications, regardless of medical care.
The virus infects roughly 90 percent of unvaccinated people exposed to it;
MAGA favorite Lauren Boebert’s son cited for child abuse once again<br /> by [[Owen Scott]] for The Independent<br /> accessed on 2026-02-20T09:36:45
A 12-Year-Old Folded Newspapers and Read Dan Bernstein's Column; Now He's Naming a Writing Room After Him<br /> by [[Justin Pardee]]<br /> accessed on 2026-02-20T09:31:07
"Typewriters are not lovers," Marshall said. "You can have more than one."
"Riverside wasn't love at first sight for me," Bernstein said. "In fact, the smog was often so thick it was hard to see the place. Walking out of the air-conditioned office and into the summer heat made me feel like I was being advised to forget about heaven."
Briefing : L'Accès au Logement des Familles Sans-Abri avec Enfants
La situation des enfants sans domicile en France atteint des seuils critiques, avec une projection de 80 000 enfants en habitat précaire pour 2025.
Malgré l'obligation légale d'hébergement d'urgence inconditionnel (Article L345-2-2 du Code de l’Action sociale), la saturation des dispositifs entraîne un tri des publics de plus en plus sévère, excluant désormais des nourrissons et des femmes enceintes.
Le recours massif à l'hébergement hôtelier, bien que palliatif, s'avère délétère pour le développement de l'enfant en raison de l'insalubrité, de l'instabilité résidentielle et d'un manque criant d'accompagnement social.
Les représentants associatifs dénoncent une « insincérité budgétaire » et une dilution des responsabilités entre l'État et les départements.
La solution identifiée réside dans un pilotage national coordonné — inspiré de la gestion de la crise ukrainienne — et une transition structurelle de l'hébergement d'urgence vers le logement social pérenne.
--------------------------------------------------------------------------------
Le constat dressé par la Fédération des acteurs de la solidarité et l'UNICEF révèle une dégradation constante :
• Chiffres globaux : 80 000 enfants sans domicile ou en habitat précaire prévus en 2025 ; 70 000 grandissent à l'hôtel ou en structures d'urgence.
• Progression du sans-abrisme : Le nombre d'enfants à la rue a augmenté de 30 % en trois ans. En octobre 2024, environ 2 500 enfants ont été recensés à la rue.
• Saturation du 115 : Environ 69 % des demandes d'hébergement restent non pourvues chaque jour. 79 % des familles ayant sollicité le 115 déclarent avoir dormi à la rue la veille de leur appel.
La loi prévoit que toute personne en détresse doit avoir accès à un hébergement à tout moment. Or, la pénurie de places force une « valse des précarités » :
• Durcissement des critères : La priorité, autrefois accordée aux enfants de moins de 3 ans et aux femmes enceintes, est désormais restreinte (femmes enceintes de plus de 6 mois, enfants de plus d'un an parfois exclus).
• Rupture de continuité : Des familles sont hébergées pour de très courtes durées (3 à 7 jours) avant d'être remises à la rue pour laisser la place à des profils jugés « plus vulnérables ».
--------------------------------------------------------------------------------
Le parc hôtelier accueille près de 30 000 enfants, dont près de 10 000 ont moins de trois ans.
Ce mode d'hébergement présente des défaillances majeures :
• Vétusté et insalubrité : Présence fréquente de nuisibles (punaises de lit, cafards).
• Absence de besoins fondamentaux : Manque d'espace, absence de cuisine (impossibilité de préparer des repas équilibrés), manque d'intimité pour les familles.
• Éloignement : Localisation souvent périphérique, rendant l'accès aux soins et aux écoles complexe (parfois des heures de transport pour maintenir la scolarité).
• Désert social : Contrairement aux centres d'hébergement d'urgence (CHU), les hôtels n'ont pas de travailleurs sociaux sur place.
• Inégalité territoriale : En Île-de-France, seules 45 % des familles à l'hôtel bénéficient d'un accompagnement via des plateformes mobiles (PASH).
• Perte de sens : Le manque de moyens réduit l'accompagnement à une variable d'ajustement budgétaire, sacrifiant l'humain au profit du paiement des nuitées.
--------------------------------------------------------------------------------
• Troubles psychiques : La prévalence des troubles de la santé mentale est de 19 % chez les enfants hébergés, contre 8 % dans la population générale.
• Insécurité permanente : Le changement constant d'environnement crée une « insécurité psychique » profonde.
• Risques sanitaires : Augmentation des naissances par césarienne (1/3 des accouchements chez les femmes sans domicile) et risques accrus de diabète gestationnel ou d'infections respiratoires.
L'instabilité résidentielle entrave l'inscription et l'assiduité scolaire.
Les enfants témoignent de l'impossibilité de nouer des liens sociaux normaux (interdiction d'inviter des amis, stigmatisation liée au lieu de vie).
• Peur du placement : Une crainte persistante des parents que la précarité du logement soit interprétée comme une défaillance éducative, entraînant un placement des enfants.
• Séparation des familles : Certains dispositifs n'acceptent que les mères et les enfants, excluant le père et brisant la cohésion familiale.
--------------------------------------------------------------------------------
Une confusion persiste sur les compétences : l'État est responsable de l'hébergement d'urgence généraliste, tandis que les Départements sont responsables de l'hébergement des mères avec enfants de moins de 3 ans.
Cette division crée des « zones grises » où des familles se retrouvent sans aucune prise en charge.
Les acteurs soulignent que les crédits alloués (785 millions d'euros pour le programme 177 en 2024) sont insuffisants pour couvrir les 203 000 places théoriques.
Cette sous-dotation structurelle empêche la création de places pérennes et de qualité.
La saturation est aggravée par la lenteur du traitement des titres de séjour (jusqu'à 18 mois d'attente).
Des personnes ayant droit au travail ou au logement social restent bloquées en hébergement d'urgence faute de documents à jour, embolisant ainsi le système.
--------------------------------------------------------------------------------
| Axe d'intervention | Actions préconisées | | --- | --- | | Pilotage | Création d'une coordination nationale interministérielle (Santé, Éducation, Logement) sur le modèle de la crise ukrainienne. | | Logement | Investissement massif dans le logement social (PLAI et PLUS) pour libérer le parc d'hébergement d'urgence. | | Qualité de l'accueil | Établissement d'un cahier des charges contraignant pour les structures accueillant des familles (accès cuisine, espaces de jeu, intimité). | | Accompagnement | Revalorisation des prix journaliers pour financer des équipes pluridisciplinaires (éducateurs de jeunes enfants, travailleurs sociaux). | | Petite Enfance | Garantie d'accès aux modes de garde (crèches) pour les familles précaires afin de favoriser la réinsertion. | | Dispositifs SAS | S'assurer que les structures d'accueil et de desserrement (SAS) garantissent une réelle évaluation et une continuité de parcours, sans retour à la rue. |
Conclusion des intervenants : L'hébergement d'urgence ne doit plus être une solution de long terme.
La priorité absolue doit être le passage direct au logement (« Logement d'abord »), seul garant de la stabilité nécessaire à la construction des enfants et à la dignité des familles.
https://www.reddit.com/r/typewriters/comments/1r98cmz/royal_qdl_used_by_jackie_robinson/

At the Jackie Robinson Museum. According to the exhibit, it was used by him to write a column for the New York Post and later the New York Amsterdam News.
Jackie Robinson used a first iteration version of Henry Dreyfuss' Royal Quiet De Luxe. The museum dates it as 1949.
Dogcatcher – Sober Me (Lyric Video)<br /> by [[Dogcatcher]] on YouTube<br /> accessed on 2026-02-20T09:00:13
Lyrics typed in real time on a typewriter as part of the music video.
via https://www.reddit.com/r/typewriters/comments/1r9pwai/lyric_video_made_with_this_subreddits_help/
The doctor was a handsome, big-shouldered man with ,1 tarnwd face.
This sentence shows how physical description helps readers quickly form an image of a character and get a sense of their presence. It shows how specific details can begin to reveal personality and set expectations about who the character might be.
Rc-1! pel1pk don't act in "general" ways
This sentence highlights the idea that writers should focus on specific, unique details instead of broad assumptions about how people behave. It emphasizes that believable characters come from showing individual reactions rather than relying on stereotypes or generalizations.
Human beings are indefinitely multifaceted and behave in ways that are seemingly contradictory.
This sentence shows that real people are complex and don’t always act in perfectly consistent ways. It explains why believable characters in fiction should show contradictions and depth rather than being simple or predictable.
Flat characters are also called stereotypes, and the hallmark of flat characters is that they are incapable of surprising us
This sentence explains that flat characters feel predictable and one dimensional because they always act the way we expect. It highlights why writers aim to create more complex, surprising characters that feel real to readers.
Welcome to Mila's technical documentation.
Hi :)
Art is intertwined with humanity, with all its flawed dimensions, and the two cannot be separated. In the making of art, there is family, there are friends and collaborators, there is both fragility and permanence, and there is the passage of time. And there is physicality.
This further shows why people didn't like this ad and how the author who wrote this has a personal attachment to music. This could mean that other musicians would be upset, but non-musicians would care a bit less.
It can replace them all. You can make your music with it. You can paint with it. You can play games on it. You can take your photos with it.
The hydrolic press materials seem to disappear after being crushed and turn into the iPad, this quote that apple implies can offend musicians because it makes music seem easy. The author of this having a connection to music could be one reason they don't like it. This quote could be interpreted that the good memories people have with music, like the author, could be replaced with an ipad. Making those experiences seem easily replaceable.
Steam engines made it possible to sail upstream as easily and quickly as down, causing an explosion of travel and shipping that radically changed frontier life.
This helped me understand how big of an impact steamboats had on travel and shipping goods. This is because they were able to go in any direction not just the direction of the current.
New technology also made mass production possible, which first occurred in the British textile industry.
Britain's industrial revolution was able to really take off because of the new machines that were getting created. It helped make it possible to produce goods in a larger amount a lot faster than before.
Links from the video: Article about 12 Favorite Problems by Joe Balcom: Article about working with email by Tiago Forte: https://fortelabs.com/blog/one-touch-... Contents: 0:00 Information. There's too much of it 2:22 What you'll learn 2:52 Principle 1 - Filtering 4:52 Principle 2 - Know the format by sight 6:11 Principle 3 - No one is forgotten, nothing is forgotten 7:45 Principle 4 - Balanced information digestion 10:22 What's the output 11:11 Finale 👋🏻Hi, I'm Max Ryzhkov. 🧘♂️I'm engaged in the integral development of myself and the systems around me through analytics, writing, and translations from complex to understandable. ☀️I love memes, I hate slush.
1233434234123
hippocampus, the entorhinal cortex, and the perihinal cortex which are important structures in the limbic system
are these parts of the brain? what do they do?
Мои заметки
eLife Assessment
This study provides valuable insights into the regulation of myogenic differentiation by identifying Leiomodin 1 as a modulator of proteome dynamics during myogenic differentiation. The combination of quantitative proteomics with functional perturbation experiments offers solid evidence supporting the idea that SIRT1 influences perturbations of myogenic differentiation upon LMOD1 inactivation. These findings advance our understanding of muscle differentiation and will be of interest to researchers studying muscle development and related pathologies
Reviewer #1 (Public review):
The main significance of this work is characterizing the function of a new gene Lmod1 in muscle stem cell biology. The study suggests an intriguing regulatory mechanism by which Sirt1 sequesters Lmod1 in a specific temporal window during myogenesis.
Comments on revisions:
The authors have satisfactorily addressed my inquires. Thank you.
Reviewer #2 (Public review):
Summary:
In this manuscript, the authors identify Leiomodin-1 (LMOD1) as a key regulator of early myogenic differentiation, demonstrating its interaction with SIRT1 to influence SIRT1's cellular localization and gene expression. The authors propose that LMOD1 translocates SIRT1 from the nucleus to the cytoplasm to permit the expression of myogenic differentiating genes such as MYOD or Myogenin.
Strengths:
A major strength of this work lies in the robust temporal resolution achieved through a time-course mass spectrometry analysis of in vitro muscle differentiation. This provides novel insights into the dynamic process of myogenic differentiation, often under explored in terms of temporal progression. The authors provide a strong mechanistic case for how LMOD1 exerts its role on muscle differentiation which opens avenues to modulate.
Weaknesses:
In the revised manuscript, the authors begin to translate their in vitro findings to an in vivo context by examining SIRT1 expression across a regeneration time course (Fig. 4I). They observe an increase in SIRT1 expression concomitant with LMOD1, supporting a potential role for SIRT1 in myogenic differentiation. Future studies will be required to provide deeper mechanistic insight into SIRT1 function in vivo.
Discussion:
Overall, the study emphasizes the importance of understanding the temporal dynamics of molecular players during myogenic differentiation and provides valuable proteomic data that will benefit the field. Future studies should explore whether LMOD1 modulates the nuclear-cytoplasmic shuttling of other transcription factors during muscle development and how these processes are mechanistically achieved. Investigating whether LMOD1 can be therapeutically targeted to enhance muscle regeneration in contexts such as exercise, aging, and disease will be critical for translational applications. Additionally, elucidating the interplay among LMOD1, LMOD2, and LMOD3 could uncover broader implications for actin cytoskeletal regulation in muscle biology. The authors have nicely updated their discussion.
Reviewer #3 (Public review):
Summary:
In this manuscript, the investigators identified LMOD1 as one of a subset of cytoskeletal proteins that levels increase in early stages of myogenic differentiation. Lmod1 is understudied in striated muscle and in particular in myogenic differentiation. Thus, this is an important study. It is also a very thorough study, with perhaps even too much data presented. Importantly, the investigators observed that LMOD1 appears to be important for skeletal regeneration, myogenic differentiation and that it interacts with SIRT1. Both primary myoblast differentiation and skeletal muscle regeneration were studied. Rescue experiments confirmed these observations: SIRT1 can rescue perturbations of myogenic differentiation as a result of LMOD1 knockdown.
Strengths:
Particular strengths include: an important topic, the use of primary skeletal cultures, the use of both cell culture and in vivo approaches, careful biomarker analysis of primary mouse myoblast differentiation, the use of two methods to probe the function of the Lmod1/SIRT1 pathway via using depletion approaches and inhibitors, and the generation of six independent myoblast cultures. Results support their conclusions.
Weaknesses:
(1) Figure 1. Images of cells in Figure 1A are too small to be meaningful (especially in comparison to the other data presented in this figure). Perhaps make graphs smaller?
(2) Line 148 "We found LMOD2 to be the most abundant Lmod in whole skeletal muscle." This is confusing since most, if not all, prior studies have shown that Lmod3 is the predominant isoform in skeletal muscle. The two papers that are cited are incorrectly cited. Clarification to resolve this discrepancy is needed.
(3) Figure 2. Immunofluorescence (IF) panels are too small to be meaningful. Perhaps the graphs could be made smaller and more space allocated for the IF panels? This issue is apparent for just about all IF panels - they are simply too small to be meaningful. Additionally, in many of the immunofluorescence figures, the colors that were used make it difficult to discern the stained cellular structures. For example, in Figure S1, orange and purple are used - they do not stand out as well as other colors that are more commonly used.
(4) There is huge variability in many experiments presented - as such, more samples appear to be required to allow for meaningful data to be obtained. For example, Figure S2. Many experimental groups, only have 3 samples - this is highly problematic - I would estimate that 5-6 would be the minimum.
(5) Ponceau S staining is often used as a loading control in this manuscript for western blots. The area/molecular weight range actually used should be specified. Not clear why in some experiments GAPDH staining is used, in other experiments Ponceau S staining is used, and in some, both are used. In some experiments the variability of total protein loaded from lane-to-lane is disconcerting. For example, in Figure S4C there appears to be more than normal variability. Can the protein assay be redone and the samples run again?
(6) Figure S3 - Lmod3 is included in the figure but no mention of it occurs in the title of the figure and/or legend.
(7) Abstract, line 25. "overexpression accelerates and improves the formation of myotubes". This is a confusing sentence. How is it improving the formation? A little more information about how they are different than developing myotubes in normal/healthy muscle would be helpful
(8) Impossible from IF figures presented to determine where Lmod1 localizes in the myocytes. Information on its subcellular localization is important. Does it localize with Lmod2 and Lmod3 at thin filament pointed ends?
Comments on revisions:
Many comments have been adequately addressed. However, some concerns remain.
Former Concern #2. The issue with the lack of detection of LMOD3 in their muscle samples is troublesome and has not been adequately resolved in the revised manuscript. It is a fact that most, if not all, studies on Lmod3 report that it is the most abundant isoform in skeletal muscle. This issue should be discussed in the manuscript. It is recognized that a different assay was utilized in this paper. The papers that are cited continue to remain incorrect. Specifically:
Tsukada et al., reports abundance of LMOD2 in cardiac muscle, not in skeletal muscle.
Nworu et al., 2015 reports on LMOD3 in skeletal muscle.
Kiss et al.,2020. While this paper reveals an important function for Lmod2 in thin filament length regulation, it is clearly shows many examples of high expression of Lmod3 in various skeletal muscles isolated from mice.
Former Concern #3. With respect to small sample numbers. Hopefully a statistical editor is available to comment. While this reviewer is happy that other assays were used to verify their data, the problem still remains that many experimental groups only have 3 samples (with high variability).
Former Concern #3. Many immunofluorescence panels are hard to evaluate because of their small size.
Author response:
The following is the authors’ response to the original reviews.
eLife Assessment
This valuable study offers insights into the role of Leiomodin-1 (LMOD1) in muscle stem cell biology, advancing our understanding of myogenic differentiation and indicating LMOD1 as a regulator of muscle regeneration, aging, and exercise adaptation. The integration of in vitro and in vivo approaches, complemented by proteomic and imaging methodologies, is solid. However, certain aspects require further attention to improve the clarity, impact, and overall significance of the work, particularly in substantiating the in vivo relevance. This work will provide a starting point that will be of value to medical biologists and biochemists working on LMOD and its variants in muscle biology.
Thank you for the positive feedback on our manuscript and the constructive criticism provided by the reviewers that helped us improve our manuscript.
Public Reviews:
Reviewer #1 (Public review):
This manuscript by Ori and colleagues investigates the role of Lmod1 in muscle stem cell activation and differentiation. The study begins with a time-course mass spectrometry analysis of primary muscle stem cells, identifying Lmod1 as a pro-myogenic candidate (Figure 1). While the initial approach is robust, the subsequent characterization lacks depth and clarity. Although the data suggest that Lmod1 promotes myogenesis, the underlying mechanisms remain vague, and key experiments are missing. Please find my comments below.
We thank the reviewer for the positive feedback on our manuscript and the helpful comments, which helped improve it.
(1) The authors mainly rely on coarse and less-established readouts such as myotube length and spherical Myh-positive cells. More comprehensive and standard analyses, such as co-staining for Pax7, MyoD, and Myogenin, would allow quantification of quiescent, activated, and differentiating stem cells in knockdown and overexpression experiments. The exact stage at which Lmod1 functions (stem cell, progenitor, or post-fusion) is unclear due to the limited depth of the analysis. Performing similar experiments on cultured single EDL fibers would add valuable insights.
We thank the reviewer for this comment. In addition to performing standard measurements such as staining for Myogenin and Myosin Heavy Chain (Figure S2H), we focused on morphological readouts, such as myotube formation, because LMOD1 is an actin cytoskeleton-associated protein. Therefore, we reasoned its function would be most directly reflected in structural changes during differentiation, rather than solely in early transcriptional markers.
Regarding the use of standard markers, we have already performed co-staining for Myogenin and Myosin Heavy Chain (MHC), which effectively quantifies early myogenic committed (Myogenin+/MHC-) and terminally differentiating (Myogenin+/MHC+) cells (Figure S2H). We did not include Pax7 as our primary culture system consists of already activated myoblasts, where Pax7 is not a reliable marker of quiescence. Our data also suggest that Lmod1 is important in regulating differentiation with comparably only mild effects on proliferation (S2D-E), therefore, we focused on this stage of myogenesis.
Our focus on differentiation over activation is further supported by multiple lines of evidence. First, analysis of publicly available transcriptome datasets reveals that Lmod1 mRNA levels actually decrease upon Muscle Stem Cell (MuSC) activation, suggesting its primary role is not during this initial phase. We added this data for clarification to Figure S1B. This aligns perfectly with our in vivo data from cardiotoxin-induced muscle regeneration, where abundance of LMOD1 protein peaks at days 4-7 post-injury — a time point coinciding with new myofiber formation and maturation — rather than during the initial activation and proliferation phase (days 1-3) (Figure 4I).
Given this strong evidence pointing to a primary role for LMOD1 during the later stages of differentiation, we believe our current analyses are the most relevant. While single EDL fiber cultures are valuable for studying the quiescence-to-activation transition, they would not provide significant additional insight into the specific differentiation-centric mechanism we are investigating here. We are confident that our chosen readouts appropriately address Lmod1's function in the differentiation of myoblasts and formation of myotubes.
(2) In supplementary Figure 2E, the distinction between Hoechst-positive cells and total cell counts is unclear. The authors should clarify why Hoechst-positive cells increase and relabel "reserve cells," as the term is confusing without reading the legend.
We thank the reviewer for pointing out the confusion regarding the naming of the cell populations and the increase in Hoechst-positive cells. We have now modified this and revised the terminology used in Figure S2E to improve clarity. Specifically, we have relabeled "reserve cells" as "non-proliferating myoblasts (Ki67-/Hoechst+)" to describe these cells more accurately without requiring the legend for interpretation. Regarding the increase in Hoechst-positive cells, we observed a slight (26%) but significant decrease in the number of proliferating myoblasts (Ki67+/Hoechst+) (Figures S2D and S2E). The relative increase in non-proliferating (Ki67-/Hoechst+) cells is a consequence of the significant reduction in the number of proliferating cells (Ki67+/Hoechst+) cells. Importantly, the total cell count (sum of Ki67-/Hoechst+) and (Ki67+/Hoechst+) remained stable. This has been clarified in the revised figure legend and main text as follows:
“This was accompanied by a proportional increase in non-proliferating myoblasts (Ki67-/Hoechst+), while the total Hoechst-positive cell count (Ki67+/Hoechst+ and Ki67-/Hoechst+) remained unchanged (Figure S2E).”
(3) The specificity of Lmod1 and Sirt1 immunostaining needs validation using siRNA-treated samples, especially as these data form the basis of the mechanistic conclusions.
We have validated the specificity of the LMOD1 antibody using multiple approaches. Specifically, we performed immunofluorescence and immunoblotting on Lmod1 siRNA-transfected samples, where we observed a significant reduction in the Lmod1 protein signal compared to control conditions (see manuscript data from Figure S2G).
Additionally, LMOD1 overexpression experiments demonstrated a corresponding increase in the signal for LMOD1 using immunofluorescence analyses, confirming the specificity of the antibody for detecting LMOD1.
For the reviewers’ interest, we add Author response image 1:
Author response image 1.
Specificity of antibodies detecting LMOD1. Representative immunofluorescence images of LMOD1 in primary myoblast cultures following siLmod1 knockdown, LMOD1 overexpression, or controls transfected with a non-targeting siRNA (siCtrl) after one day of differentiation. LMOD1 (purple), SIRT1 (yellow), and nuclei (Hoechst, blue). Scale bar: 10 µm.
For the SIRT1 antibody used in our immunostaining, the specificity was validated by transfecting primary myoblasts with siRNA targeting Sirt1 and performing immunoblot analyses (Figure S5A). These showed a significant reduction in SIRT1 protein levels, confirming both the effectiveness of the siRNA and, critically, the antibody's ability to specifically recognize and detect SIRT1 protein. Furthermore, the same SIRT1 antibody was utilized in our nuclear-cytoplasmic fractionation experiments (Figure S4C), and its ability to detect SIRT1 in the expected subcellular compartments further supports its specific binding to SIRT1. While direct immunofluorescence on Sirt1 siRNA-transfected samples was not performed, the robust demonstration of the antibody's specificity for Sirt1 protein via immunoblotting (i.e., correct molecular weight band, significantly reduced by Sirt1 siRNA) and its distribution in subcellular fractions, which is fully consistent with the localization immunostaining performed at the same time points (compare Figure S4C and 5A), provide strong evidence on the antibody’s specificity, also in immunofluorescence experiments.
(4) The authors must test the effect of Lmod1 siRNA on Sirt1 localization, as only overexpression experiments are shown
We carefully considered performing this experiment. However, the knockdown of Lmod1 significantly impairs myogenic differentiation, a crucial cellular process that itself can influence protein localization. Consequently, if SIRT1 localization would be altered following knockdown of Lmod1, it would be challenging to disentangle whether this was a direct result of LMOD1 absence impacting SIRT1 trafficking or an indirect consequence of the cells failing to differentiate properly. This would make it difficult to draw clear conclusions regarding a direct causal link between LMOD1 and SIRT1 localization from such an experiment. Therefore, we focused on overexpression experiments, where we could demonstrate that altering LMOD1 levels is sufficient to affect SIRT1 localization. Our nuclear-cytoplasmic fractionation experiments clearly show that LMOD1 overexpression leads to changes in SIRT1 distribution (Figure 5H-K). These findings provide evidence that LMOD1 can directly modulate SIRT1 localization, supporting our mechanistic conclusions.
(5) In Figure S3, the biotin signal in LMOD2 samples appears weak. The authors need to address whether comparing LMOD1 and LMOD2 is valid given the apparent difference in reaction efficiency. It would also help to highlight where Sirt1 falls on the volcano plot in S3B.
We agree that the overall biotin signal on the streptavidin blot for the LMOD2-BirA* sample appears weaker than for LMOD1-BirA*. To provide a more direct comparison of the bait proteins themselves, we have now added a bar graph to the revised Figure S3D, which quantifies the relative abundance of LMOD1 and LMOD2 bait proteins in the pull down experiments. This analysis shows that the levels of LMOD1-BirA* and LMOD2-BirA* were comparable in our BioID samples. Furthermore, the validity of the LMOD2 BioID experiment is strongly supported by the identification of several known LMOD1 and LMOD2 interaction partners. As shown in the dataset, well-established interactors such as TMOD1, TPM3, and TMOD3 were identified, with some even showing stronger enrichment with LMOD2 than with LMOD1. This confirms that the biotinylation reaction was efficient enough to capture proximal proteins for both baits.
Regarding SIRT1, we have now highlighted in yellow its position on the volcano plot in the revised Figure S3E. As can be seen, SIRT1 was identified in the LMOD1-BirA sample and showed enrichment. We believe these clarifications, along with the additional expression data and the successful identification of known interactors, confirm the validity of our comparative BioID analysis.
(6) The immunostaining data suggest that Lmod1 remains cytoplasmic throughout differentiation, whereas Sirt1 shows transient cytoplasmic localization at day 1 of differentiation. The authors should explain why Sirt1 is not constantly sequestered if Lmod1's cytoplasmic localization is consistent. It is also unclear whether day 1 is the key time point for Lmod1 function, as its precise role during myogenesis remains ambiguous.
We thank the reviewer for this comment. We have no data explaining why SIRT1 is not constantly sequestered while LMOD1 remains consistently cytoplasmic. We can only speculate that the transient cytoplasmic localization of SIRT1 may be linked to the availability and functional role of LMOD1 throughout the differentiation process. While LMOD1 is present at low levels in proliferating primary myoblasts, its expression increases upon the initiation of differentiation (Figure 2A). Initially, during the early stages of differentiation, LMOD1 may not be required for actin nucleation as the major remodeling of the cytoskeleton has not yet begun. During this phase, LMOD1 might have the capacity to sequester SIRT1 in the cytoplasm.
However, as differentiation progresses and morphological changes take place, LMOD1 may switch its functional role to actin nucleation, thereby releasing SIRT1. This transition could explain why SIRT1 is free to localize transiently to the cytoplasm, particularly at day 1, when cytoskeletal remodeling is beginning but not yet fully established.
Additionally, as LMOD1 and SIRT1 are known to colocalize in the nucleus, they may exit the nucleus together. Once in the cytoplasm, LMOD1 may become engaged in actin nucleation, allowing SIRT1 to function independently, which could explain the transient nature of SIRT1’s cytoplasmic localization.
We have acknowledged this gap in our understanding in the discussion of the revised manuscript:
“Our immunostaining data show that while LMOD1 is consistently cytoplasmic, its partner SIRT1 is only transiently localized in the cytoplasm. This suggests that their interaction is dynamically regulated. We hypothesize that the function of LMOD1 is determined by the changing availability of its binding partners during differentiation. During the initial phase, LMOD1 may primarily function to sequester SIRT1, a key regulator of myogenic genes. As differentiation proceeds, the increased expression of cytoskeletal components, such as its canonical partners TMODs and TPMs, likely shifts the function of LMOD1 towards its role in actin nucleation. This molecular switch, potentially driven by a change in the interactome of LMOD1, could then result in the release of SIRT1 from the cytoplasm. Such a mechanism may coordinate transcriptional regulation with cytoskeletal remodeling during myoblast differentiation.”
(7) The introduction does not sufficiently establish the motivation or knowledge gap this work aims to address. Instead, it reads like a narration of disparate topics in a single paragraph. The authors should clarify the statement in line 150, "since this protein has been...,".
We thank the reviewer for requesting clarification regarding our focus on LMOD1 (Introduction and Line 150 in the original submission). In the revised manuscript, we shortened the introduction and more clearly emphasized the motivation of our study:
“Although these mechanisms contribute to remodeling the cellular architecture of MuSCs, a comprehensive understanding of the temporal dynamics of proteome remodeling during differentiation remains lacking. To address this knowledge gap, we performed an unbiased proteomic analysis of the early stages of myogenic differentiation to identify previously unrecognized proteins involved in this process and to examine how they functionally interact with established regulatory pathways.”
Our decision to focus on LMOD1 was driven by its significant upregulation in our temporal proteome dataset, together with its previously uncharacterized role in primary myoblasts. Furthermore, to strengthen the interpretation of LMOD1’s role, particularly in the context of aging, we have integrated a new analysis of published transcriptomic datasets. This can be found in the main text as follows:
“Surprisingly, we detected LMOD1 in freshly isolated muscle stem cells (MuSCs), but not LMOD2. Additionally, we observed that the protein levels of LMOD1 increased in MuSCs isolated from older mice (Figure 2C and Figure S1B). We further analyzed published transcriptomic data sets that describe changes between young and old MuSCs in both quiescent and activated states in young and old animals (Liu et al. 2013; Lukjanenko et al. 2016). In these analyzed transcriptomic data sets, Lmod1 was found to be significantly downregulated during the activation of MuSCs in both young and old mice (see Figure S1B).
To assess the in vivo relevance of our finding, we queried two proteomic datasets of freshly isolated MuSCs and four different skeletal muscles (gastrocnemius, G; soleus, S; tibialis anterior, TA; extensor digitorum longus, EDL) (Schüler et al. 2021). We found LMOD2 to be the most abundant leiomodin protein in whole skeletal muscle, consistent with data from (Tsukada et al. 2010; Nworu et al. 2015; Kiss et al. 2020), while the overall abundance of LMOD1 was lower since this protein has been mainly associated with smooth muscle cells (Nanda and Miano 2012; Conley et al. 2001; Nanda et al. 2018) (Figure 2B).”
Overall, while the identification of Lmod1 as a pro-myogenic factor is convincing, the mechanistic insights are insufficient, and the manuscript would benefit from addressing these concerns.
We thank the reviewer for their constructive criticism. In the revised manuscript, we have strengthened our mechanistic insights and the validation of our findings by implementing the suggestions of the reviewers and including new experimental data to address their concerns.
Reviewer #2 (Public review):
Summary:
In this manuscript, the authors identify Leiomodin-1 (LMOD1) as a key regulator of early myogenic differentiation, demonstrating its interaction with SIRT1 to influence SIRT1's cellular localization and gene expression. The authors propose that LMOD1 translocates SIRT1 from the nucleus to the cytoplasm to permit the expression of myogenic differentiation genes such as MYOD or Myogenin.
Strengths:
A major strength of this work lies in the robust temporal resolution achieved through a time-course mass spectrometry analysis of in vitro muscle differentiation. This provides novel insights into the dynamic process of myogenic differentiation, often under-explored in terms of temporal progression. The authors provide a strong mechanistic case for how LMOD1 exerts its role in muscle differentiation which opens avenues to modulate.
We thank the reviewer for the positive feedback on our manuscript and the insightful comments which helped to improve the manuscript!
Weaknesses:
One limitation of the study is the in vivo data. Although the authors do translate their findings in vivo for LMOD1 localization and expression, the cross-sectional imaging is not highly convincing. Longitudinal cuts or isolated fibers could have been more useful specimens to answer these questions. Moreover, the authors do not assess their in vitro SIRT1 findings in vivo. A few key experiments in regenerating or aged mice would strengthen the mechanistic insight of the findings.
We agree that longitudinal cuts and isolated fibers can provide excellent morphological detail for specific questions. However, for our primary objective in this study, which was to assess the temporal expression and localization of LMOD1 across the tissue during the regeneration process, we decided that cross-sectional analysis provided the most robust and reliable overview. Cross-sectional imaging effectively captures the spatial distribution of LMOD1 across multiple myofibers and their surrounding microenvironment, simultaneously assessing the whole cross-sectional area. By using this approach, we were able to evaluate the broader tissue architecture and cellular context, which was essential for understanding the dynamic changes occurring during regeneration. We were also able to investigate all myofibers of a muscle, and not only a small proportion, which we would analyze with longitudinal sections and isolated myofibers. Therefore, we continued using cross-sections for further analyses.
We fully agree with the reviewer that validating our in vitro SIRT1 findings in an in vivo context is an essential next step. To address this, we performed additional analyses on our existing regenerating muscle samples and incorporated new immunostainings for SIRT1 and PAX7 into the regeneration time-course (now shown in revised Figure 4I), providing further in vivo support for our proposed mechanism. We focused specifically on cross-sections collected at day 5 post-injury, a time point selected based on the peak in LMOD1 expression, to assess whether SIRT1 levels increase in parallel with LMOD1 during regeneration. Notably, SIRT1 abundance is elevated at day 5 post-injury, underscoring its involvement in early myogenic differentiation. This conclusion is further supported by the localization of SIRT1 within mononucleated cells and newly formed myofibers at this stage of regeneration.
Finally, we agree that further mechanistic studies in vivo would be highly valuable. While we were able to address SIRT1 dynamics in our regeneration model as suggested, an aged mouse cohort was unfortunately not available to us for this kind of study. Furthermore, more extensive in vivo experiments, such as those involving genetic manipulation, were beyond the scope of the current study, partly due to constraints related to animal welfare regulations and our approved experimental protocols.
Discussion:
Overall, the study emphasizes the importance of understanding the temporal dynamics of molecular players during myogenic differentiation and provides valuable proteomic data that will benefit the field. Future studies should explore whether LMOD1 modulates the nuclear-cytoplasmic shuttling of other transcription factors during muscle development and how these processes are mechanistically achieved. Investigating whether LMOD1 can be therapeutically targeted to enhance muscle regeneration in contexts such as exercise, aging, and disease will be critical for translational applications. Additionally, elucidating the interplay among LMOD1, LMOD2, and LMOD3 could uncover broader implications for actin cytoskeletal regulation in muscle biology.
We thank the reviewer for this excellent suggestion for future analyses. We have included these important considerations and future avenues in the Discussion of the revised manuscript:
“Our immunostaining data show that while LMOD1 is consistently cytoplasmic, its partner SIRT1 is only transiently localized in the cytoplasm. This suggests that their interaction is dynamically regulated. We hypothesize that the function of LMOD1 is determined by the changing availability of its binding partners during differentiation. During the initial phase, LMOD1 may primarily function to sequester SIRT1, a key regulator of myogenic genes. As differentiation proceeds, the increased expression of cytoskeletal components, such as its canonical partners TMODs and TPMs, likely shifts the function of LMOD1 towards its role in actin nucleation. This molecular switch, potentially driven by a change in the interactome of LMOD1, could then result in the release of SIRT1 from the cytoplasm. Such a mechanism may coordinate transcriptional regulation with cytoskeletal remodeling during myoblast differentiation.”
“Moreover, delineating the functional specialization and potential redundancy among leiomodin proteins represents an important next step. Our data indicate that LMOD1 primarily regulates early myogenic differentiation (Figure 3). In contrast, the lack of an early functional phenotype upon LMOD2 depletion, together with its upregulation at later stages (Figure S2A), suggests a temporal shift in regulatory control. Accordingly, a systematic comparative analysis of LMOD1, LMOD2, and LMOD3 will be required to elucidate their distinct roles in actin cytoskeleton regulation across the myogenic program, particularly with respect to myofibril maturation and maintenance.”
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
Major Changes:
(1) In Vivo Data on SIRT1:
The inclusion of in vivo data on SIRT1 localization and expression would significantly strengthen the manuscript. Similar staining techniques used for LMOD1 could be applied to SIRT1. Additionally, imaging muscle specimens such as longitudinal sections or isolated myofibers would provide clearer insights into SIRT1's spatial distribution and improve upon the less convincing cross-sectional images currently presented (Figure 2).
We fully agree that providing in vivo data on SIRT1 localization and expression is a crucial step to support our in vitro findings. Following the reviewer's suggestion, we have performed new experiments on muscle regeneration samples using the analyses of cross-sections as done for the analysis of LMOD1 localization. Specifically, we performed immunostaining for SIRT1 on cross-sections from muscle samples collected at day 5 post-injury, a time point selected based on the observed peak in LMOD1 expression. These new data (now included in revised Figure 4I) allowed us to assess whether SIRT1 levels increase during regeneration in parallel with an increase in LMOD1 abundance.
Regarding the suggestion to use longitudinal sections or isolated myofibers, we agree that these preparations offer excellent answers for certain questions. For the primary goal of our study, to assess the temporal expression changes across the entire regenerating tissue at different time points, we found that cross-sections provided the most comprehensive and robust overview and therefore did not use longitudinal sections or isolated myofibers.
Performing additional animal experiments to obtain these specific preparations was beyond the scope of the current study and subject to constraints from our approved animal welfare protocols.
(2) Morphology of siLmod1 Cells:
The morphology of siLmod1-treated cells in vitro (Figure 3) raises concerns. Assessing cell viability or cell death in these experiments would help ensure that differences are not due to dead or unhealthy cells being quantified. There is also a notable discrepancy between the control panels in Figures 3C and 3H compared to the experimental conditions in 3F and 3K, particularly in terms of cell length and morphology. These inconsistencies should be addressed or clarified.
We acknowledge the visual discrepancies in cell morphology noted by the reviewer (e.g., between Figures 3C/3H and 3F/3K). These differences can be attributed to biological variability between primary myoblast cultures isolated from different mice. Such variability includes differences in myogenic potential and the fact that cells are not synchronized, leading to variations in differentiation efficiency, baseline morphology, and cell length across cultures (Cornelison 2008; Vaughan and Lamia 2019). To account for this, we decided to use n=6 biological replicates, i.e., primary myoblast cultures isolated from 6 different mice, for immunofluorescence analysis, ensuring robust quantitative data. Furthermore, we confirmed that this phenotype was not an artifact of culture conditions, as we consistently observed the same effect of Lmod1 knockdown independently of the passage number of the myoblasts or the donor mouse.
To address the concerns that morphological changes in siLmod1-treated cells might reflect cell death, we performed a TUNEL assay (transfection at day 1, analysis at day 3 of differentiation). This revealed no significant increase in TUNEL-positive (apoptotic) cells in siLmod1- (or siSirt1-) transfected samples versus siCtrl-transfected cells. These new data have been added to the revised manuscript as Supplementary Figure S2I. The TUNEL data indicate that the observed morphological changes upon knockdown of Lmod1 are not due to induced cell death. Supported by these results, our interpretation is that knockdown of Lmod1 impairs or arrests differentiation rather than causing cell death. Furthermore, our quantification of different cell populations showed shifts indicative of impaired differentiation (e.g., accumulation of cells at earlier stages) without exhibiting significant loss in cell numbers. For example, the numbers of myogenin+/MHC- and myogenin+/MHC+ cell populations, and differentiated myotubes, were not significantly reduced after transfection with siLmod1. A slight, not significant trend towards fewer non-proliferating myoblasts/reserve cells characterized by the expression of Myogenin-/MHC-Hoechst+ (Figure S2H) was noted. Overall, cells appeared to be 'stuck' in differentiation, consistent with the role of Lmod1 in impairing differentiation but not causing cell death. We have further clarified this aspect in the revised manuscript.
(3) LMOD1 and SIRT1 Interaction in Myogenic Cells:
Strengthening the connection between LMOD1 and SIRT1 within the myogenic system would enhance the manuscript. Could proximity ligation assays (PLA) be performed in myogenic cells, as was done in HEK293T cells? Additionally, investigating whether SIRT1 remains in the nucleus upon LMOD1 knockdown using siRNA would provide mechanistic insight into their interaction during myogenic differentiation.
We would like to clarify that the Proximity Ligation Assays (PLA) shown in Figure 4H were indeed performed in primary myoblasts, confirming the LMOD1-SIRT1 interaction directly in a myogenic context. We have modified the text to clarify that primary myoblasts were used for the PLA assays.
Minor Points:
(1) Was Lmod1 knockdown confirmed in vivo?
To target Lmod1 in Muscle Stem Cells (MuSCs) in vivo, we utilized self-delivering Accell siRNAs. This delivery system has been previously validated and shown to be highly effective for targeting MuSCs in regenerating muscle (Bentzinger et al., Cell Stem Cell, 2013).
While this is an established method for delivery, confirming knockdown specifically within the rare MuSC population is technically challenging using bulk tissue analysis, as the target signal is diluted by numerous other cell types.
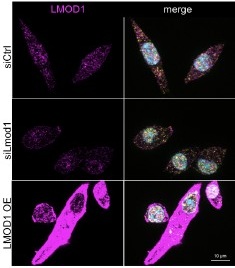
Therefore, to ensure the efficacy of our specific siRNA, we performed in vitro validation. For the reviewers' interest, we add Author response image 2 showing the efficiency of the respective siRNAs:
Author response image 2.
Knockdown efficiency of siRNAs targeting Lmod1 and Lmod2 following using the same self-delivering siRNA in proliferating primary myoblasts as used in in vivo experiments. Self-delivering Accell siRNA was added to primary myoblasts cultured in low serum media for 48 hours. Relative mRNA expression levels of Lmod1 and Lmod2 were measured after self-delivering Accell siRNA transfection targeting either Lmod1 (siLmod1) or Lmod2 (siLmod2). Expression levels were compared to control siRNA-transfected cells (siCtrl) and normalized to Gapdh expression.
Based on the documented efficacy of this delivery system from prior literature and our own validation of the specific siRNAs used here, we are confident in the knockdown efficiency of the respective siRNAs. We decided not to perform additional animal experiments due to animal welfare considerations.
(2) Some of the western blot bands do not appear to match the expected patterns for the tested proteins compared to controls (e.g., Figure S2J, S4C). Ensure that these are accurately labeled and include the entire membrane for transparency and reproducibility.
Regarding Figure S2J, we agree that the presentation could be confusing to the reader. The blot shows LMOD1 and LMOD2 knockdown, while the bar plot quantifies only the change in LMOD2 levels. We have now revised the figure legend to explicitly state this. We hope this makes the presentation of our data clearer.
For Figure S4C, we believe the concern about 'patterns' relates to loading variability. In this experiment, we manually counted the nuclei before lysis to ensure that each nuclear fraction started with an equal amount of material. We then loaded the cytoplasmic fractions in proportion to these counts. The purity of the fractions was additionally confirmed using nuclear (H4) and cytoplasmic (ALDOA) markers. As stated in the figure, the nuclear/cytoplasmic ratio of LMOD1 or SIRT1 was normalized across the entire lane of the Ponceau S staining, which we have now clarified in the relevant figure legends.
Finally, regarding transparency, the presented immunoblot images are representative crops, which is standard practice for clarity. We are committed to reproducibility and will provide full, uncropped scans of all blots in the final version of the manuscript, in line with eLife publishing guidelines.
(3) Figure S1B appears to reuse images from Figure 2D (rotated). Verify that this is acceptable for the journal's guidelines, and if necessary, provide additional justification or clarification.
We acknowledge that the image presented in Figure S1B was accidentally reused as a representative example in Figure 2D. To address this and prevent any potential redundancy or confusion, we have revised Figure S1B by replacing the duplicated image with a different, representative example from our dataset. The updated figure now contains unique image data, and we believe this revision fully resolves the concern.
(4) Ensure consistent scale bars across images, particularly in Figures 3C and 3H, where discrepancies might affect interpretation.
We thank the reviewer for pointing this out, we have now standardized all scale bars throughout the manuscript to ensure consistency. All immunofluorescence images of cultured cells (including Fig 3C and 3H) now have a 50 µm scale bar, and all tissue cross-sections have a 100 µm scale bar. This change has been implemented in the revised figures.
Reviewer #3 (Public review):
Summary:
In this manuscript, the investigators identified LMOD1 as one of a subset of cytoskeletal proteins whose levels increase in the early stages of myogenic differentiation. Lmod1 is understudied in striated muscle and in particular in myogenic differentiation. Thus, this is an important study. It is also a very thorough study - with perhaps even too much data presented. Importantly, the investigators observed that LMOD1 appears to be important for skeletal regeneration, and myogenic differentiation and that it interacts with SIRT1. Both primary myoblast differentiation and skeletal muscle regeneration were studied. Rescue experiments confirmed these observations: SIRT1 can rescue perturbations of myogenic differentiation as a result of LMOD1 knockdown.
Strengths:
Particular strengths include: important topic, the use of primary skeletal cultures, the use of both cell culture and in vivo approaches, careful biomarker analysis of primary mouse myoblast differentiation, the use of two methods to probe the function of the Lmod1/SIRT1 pathway via using depletion approaches and inhibitors, and generation of six independent myoblast cultures. Results support their conclusions.
We thank the reviewer for the positive assessment of our work and the helpful comments for improving our manuscript.
Weaknesses:
(1) Figure 1. Images of cells in Figure 1A are too small to be meaningful (especially in comparison to the other data presented in this figure). Perhaps the authors could make graphs smaller?
We have adjusted the size of the images across all figure panels to ensure better visibility and clarity. We hope these adjustments improve the presentation of the data.
(2) Line 148 "We found LMOD2 to be the most abundant Lmod in the whole skeletal muscle." This is confusing since most, if not all, prior studies have shown that Lmod3 is the predominant isoform in skeletal muscle. The two papers that are cited are incorrectly cited. Clarification to resolve this discrepancy is needed.
We acknowledge that LMOD2 and LMOD3 are predominantly expressed in skeletal and cardiac muscles (Tsukada et al. 2010; Nworu et al. 2015), www.proteinatlas.org) and LMOD3’s transcription is directly regulated by MRTF/SRF and MEF2 to coordinate sarcomeric assembly (Cenik et al. 2015). However, our statement refers specifically to the analysis of the proteomic datasets from freshly isolated MuSCs and four distinct skeletal muscles (G, S, TA, EDL) generated by Schüler et al. 2021. Crucially, LMOD3 was not detected in the quantitative mass spectrometry data for the EDL, G, S, or TA muscle samples analyzed in this specific study. In the context of this particular dataset, LMOD2 was the most highly abundant Leiomodin isoform detected in the whole skeletal muscle samples. This finding suggests a differential expression and function between LMOD isoforms depending on the muscle type and/or developmental/regenerative state. We have revised and corrected this clarification in the manuscript, including correcting the initial citations.
(3) Figure 2. Immunoflorescence (IF) panels are too small to be meaningful. Perhaps the graphs could be made smaller and more space allocated for the IF panels? This issue is apparent for just about all IF panels - they are simply too small to be meaningful. Additionally, in many of the immunofluorescence figures, the colors that were used make it difficult to discern the stained cellular structures. For example, in Figure S1, orange and purple are used - they do not stand out as well as other colors that are more commonly used.
We agree that the IF panels were too small for optimal interpretation and have adjusted them in Figure 2 and throughout the manuscript. Regarding the color choices, we appreciate the reviewer's comments. Our initial selection (e.g., orange and purple in Figure S1) was intended to enhance accessibility for individuals with common color vision deficiencies, including red-green color blindness. However, we acknowledge the reviewer's point that these combinations provided insufficient contrast for discerning cellular structures. Therefore, we have revised the color schemes to use green, red, and blue, which should offer improved contrast.
(4) There is huge variability in many experiments presented - as such, more samples appear to be required to allow for meaningful data to be obtained. For example, Figure S2. Many experimental groups, only have 3 samples - this is highly problematic - I would estimate that 5-6 would be the minimum.
We thank the reviewer for the comment regarding experimental variability and sample size. In our study, n=3 biological replicates, i.e., independent primary cell cultures obtained from different mice, were primarily used for immunoblots. We acknowledge that variability can be observed between distinct primary cell cultures due to factors such as inherent differences in myogenic potential, cell cycle state (as cells were not synchronized), and passage number. Importantly, despite this inter-sample variation, the investigated phenotypes showed consistent trends across biological replicates. Rather than increasing the number of replicates for immunoblots, we opted for validating our key findings using independent approaches with a higher number of replicates. For instance, qRT-PCR analyses (to confirm knockdown efficiency) and immunofluorescence analyses were mostly performed using five to six independent myoblast cultures (biological replicates).
(5) Ponceau S staining is often used as a loading control in this manuscript for western blots. The area/molecular weight range actually used should be specified. Not clear why in some experiments GAPDH staining is used, in other experiments Ponceau S staining is used, and in some, both are used. In some experiments, the variability of total protein loaded from lane to lane is disconcerting. For example, in Figure S4C there appears to be more than normal variability. Can the protein assay be redone and samples run again?
We have clarified in the relevant figure legends that Ponceau S normalization, when used, was based on the quantification of the entire lane. Our standard loading control is GAPDH. We used Ponceau S for normalization only when GAPDH was deemed unsuitable, e.g., in nuclear-cytoplasmic fractionation experiments where GAPDH is not present in all fractions.
Concerning the variability observed in Figure S4C, we manually counted the nuclei before lysis to ensure that each nuclear fraction started with an equal amount of material. We then loaded the cytoplasmic fractions in proportion to these counts. The purity of the fractions was additionally confirmed using nuclear (H4) and cytoplasmic (ALDOA) markers. The nuclear/cytoplasmic ratio of LMOD1 or SIRT1 was normalized across the entire lane of the Ponceau S staining, which we have now clarified in the relevant figure legends.
(6) Figure S3 - Lmod3 is included in the figure but no mention of it occurs in the title of the figure and/or legend.
We wish to clarify that the protein identified in Figure S3 is TMOD3 (Tropomodulin 3), not LMOD3. TMOD3 is a known pointed-end capping protein regulating the actin filament nucleation process together with LMODs (Fowler and Dominguez 2017; Boczkowska et al. 2015), so its presence in our dataset was expected and helps validate our results.
(7) Abstract, line 25. "overexpression accelerates and improves the formation of myotubes". This is a confusing sentence. How is it improving the formation? A little more information about how they are different than developing myotubes in normal/healthy muscles would be helpful.
We thank the reviewer for the comment. To clarify, we have revised the sentence in line 25 to "improves the initiation of myotube formation." This change reflects our observation that overexpression of LMOD1 leads to a more rapid onset of myotube formation, as evidenced by earlier expression of differentiation markers and accelerated fusion of myoblasts into myotubes compared to GFP overexpression myoblast cell line. These findings suggest that LMOD1 overexpression enhances the efficiency of the early stages of differentiation and fusion, thereby contributing to improved initiation of myotube formation.
(8) It is impossible from the IF figures presented to determine where Lmod1 localizes in the myocytes. Information on its subcellular localization is important. Does it localize with Lmod2 and Lmod3 at thin filament pointed ends?
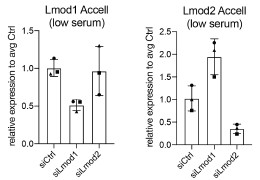
Several publications suggest that LMODs are involved in actin nucleation and interact with TMODs at the thin filament pointed ends (Boczkowska et al. 2015; Fowler and Dominguez 2017; Fowler, Greenfield, and Moyer 2003; Tsukada et al. 2010; Rao, Madasu, and Dominguez 2014). We performed F-actin (Phalloidin) staining together with LMOD1 staining and observed possible co-localization (see Author response image 3). Specifically, we noted an accumulation of LMOD1 at the ends of myocytes, indicating that LMOD1 might play a role in the elongation and guidance of myotube differentiation. For the reviewer’s interest, we include Author response image 3 as it was not part of the original manuscript. While performing subcellular localization stainings, we added the F-actin/Phalloidin staining to explore potential interactions, but this aspect was not further investigated in the current study.
Author response image 3.
Co-staining of LMOD1 and Phalloidin in differentiating myocytes.Example image showing immunofluorescence staining of LMOD1 (purple) and F-actin (green; Phalloidin) in differentiating primary myocytes. LMOD1 appears to accumulate at the ends of elongated myocytes and co-localizes with actin structures (highlighted in boxes), suggesting a potential role in myotube elongation and guidance during differentiation.
Our study focused on a distinct role for LMOD1, independent from its function in actin filament nucleation, and we therefore did not pursue further co-localization staining with LMOD2 or LMOD3. We recognize the potential importance of exploring these interactions and their relevance to thin filament organization in skeletal muscle. However, although this was beyond the scope of our current work, we will investigate this aspect in the future.
References
Boczkowska, Malgorzata, Grzegorz Rebowski, Elena Kremneva, Pekka Lappalainen, and Roberto Dominguez. 2015. “How Leiomodin and Tropomodulin Use a Common Fold for Different Actin Assembly Functions.” Nature Communications 6 (1): 8314.
Cenik, Bercin K., Ankit Garg, John R. McAnally, John M. Shelton, James A. Richardson, Rhonda Bassel-Duby, Eric N. Olson, and Ning Liu. 2015. “Severe Myopathy in Mice Lacking the MEF2/SRF-Dependent Gene Leiomodin-3.” The Journal of Clinical Investigation 125 (4): 1569–78.
Cornelison, D. D. W. 2008. “Context Matters: In Vivo and in Vitro Influences on Muscle Satellite Cell Activity.” Journal of Cellular Biochemistry 105 (3): 663–69.
Fowler, Velia M., and Roberto Dominguez. 2017. “Tropomodulins and Leiomodins: Actin Pointed End Caps and Nucleators in Muscles.” Biophysical Journal 112 (9): 1742–60.
Fowler, Velia M., Norma J. Greenfield, and Jeannette Moyer. 2003. “Tropomodulin Contains Two Actin Filament Pointed End-Capping Domains.” The Journal of Biological Chemistry 278 (41): 40000–9.
Liu, Ling, Tom H. Cheung, Gregory W. Charville, Bernadette Marie Ceniza Hurgo, Tripp Leavitt, Johnathan Shih, Anne Brunet, and Thomas A. Rando. 2013. “Chromatin Modifications as Determinants of Muscle Stem Cell Quiescence and Chronological Aging.” Cell Reports 4 (1): 189–204.
Lukjanenko, Laura, M. Juliane Jung, Nagabhooshan Hegde, Claire Perruisseau-Carrier, Eugenia Migliavacca, Michelle Rozo, Sonia Karaz, et al. 2016. “Loss of Fibronectin from the Aged Stem Cell Niche Affects the Regenerative Capacity of Skeletal Muscle in Mice.” Nature Medicine 22 (8): 897–905.
Nworu, Chinedu U., Robert Kraft, Daniel C. Schnurr, Carol C. Gregorio, and Paul A. Krieg. 2015. “Leiomodin 3 and Tropomodulin 4 Have Overlapping Functions during Skeletal Myofibrillogenesis.” Journal of Cell Science 128 (2): 239–50.
Rao, Jampani Nageswara, Yadaiah Madasu, and Roberto Dominguez. 2014. “Mechanism of Actin Filament Pointed-End Capping by Tropomodulin.” Science 345 (6195): 463–67.
Schüler, Svenja C., Joanna M. Kirkpatrick, Manuel Schmidt, Deolinda Santinha, Philipp Koch, Simone Di Sanzo, Emilio Cirri, Martin Hemberg, Alessandro Ori, and Julia von Maltzahn. 2021. “Extensive Remodeling of the Extracellular Matrix during Aging Contributes to Age-Dependent Impairments of Muscle Stem Cell Functionality.” Cell Reports 35 (10): 109223.
Tsukada, Takehiro, Christopher T. Pappas, Natalia Moroz, Parker B. Antin, Alla S. Kostyukova, and Carol C. Gregorio. 2010. “Leiomodin-2 Is an Antagonist of Tropomodulin-1 at the Pointed End of the Thin Filaments in Cardiac Muscle.” Journal of Cell Science 123 (Pt 18): 3136–45.
Vaughan, Megan, and Katja A. Lamia. 2019. “Isolation and Differentiation of Primary Myoblasts from Mouse Skeletal Muscle Explants.” Journal of Visualized Experiments: JoVE, no. 152 (October). https://doi.org/10.3791/60310.
à profissionalização
NAO CONFUNDA COM O TRABALHO
Test
eLife Assessment
This elegant study presents a valuable approach to probing the structural features of the full-length human Hv1 channel as a purified protein, supported by rigorous biochemical assays and spectral FRET analysis, which will interest biophysicists and physiologists studying Hv1 and other ion channels. Overall, the work introduces an interesting labeling strategy and provides methodological observations that are of value in investigating hHV1. However, the analysis appears incomplete, requiring additional structural interpretation and mechanistic insight.
Reviewer #1 (Public review):
In this study, the noncanonical amino acid acridon-2-ylalanine (Acd) was inserted at various positions within the human Hv1 protein using a genetic code expansion approach. The purified mutants with incorporated fluorophore were shown to be functional using a proton flux assay in proteoliposomes. FRET between native tryptophan and tyrosine residues and Acd were quantified using spectral FRET analysis. Predicted FRET efficiencies calculated from an AlphaFold model of the Hv1 dimer were compared to the corresponding experimental values. Spectral FRET analysis was also used to test whether structural rearrangements caused by Zn2+, a well-known Hv1 inhibitor, could be detected. The experimental data provide a good validation of the approach, but further expansion of the analysis will be necessary to differentiate between intra- and intersubunit structural features.
Interestingly, the observed rearrangements induced by Zn2+ were not limited to the protein region proximal to the extracellular binding site but extended to the intracellular side of the channel. This finding agrees with previous studies showing that some extracellular Hv1 inhibitors, such as Zn2+ or AGAP/W38F, can cause long-range structural changes propagating to the intracellular vestibule of the channel (De La Rosa et al. J. Gen. Physiol. 2018, and Tang et al. Brit J. Pharm 2020). The authors should consider adding these references.
Since one of the main goals of this work was to validate Acd incorporation and the spectral FRET analysis approach to detect conformational changes in hHv1 in preparation for future studies, the authors should consider removing one subunit from their dimer model, recalculating FRET efficiencies for the monomer, and comparing the predicted values to the experimental FRET data. This comparison could support the idea that the reported FRET measurements can inform not only on intrasubunit structural features but also on subunit organization.
Reviewer #2 (Public review):
This manuscript by Carmona, Zagotta, and Gordon is generally well-written. It presents a crude and incomplete structural analysis of the voltage-gated proton channel based on measured FRET distances. The primary experimental approach is Förster Resonance Energy Transfer (FRET), using a fluorescent probe attached to a noncanonical amino acid. This strategy is advantageous because the noncanonical amino acid likely occupies less space than conventional labels, allowing more effective incorporation into the channel structure.
Fourteen individual positions within the channel were mutated for site-specific labeling, twelve of which yielded functional protein expression. These twelve labeling sites span discrete regions of the channel, including P1, P2, S0, S1, S2, S3, S4, and the dimer-connecting coiled-coil domain. FRET measurements are achieved using acridon-2-ylalanine (Acd) as the acceptor, with four tryptophan or four tyrosine residues per monomer serving as donors. In addition to estimating distances from FRET efficiency, the authors analyze full FRET spectra and investigate fluorescence lifetimes on the nanosecond timescale.
Despite these strengths, the manuscript does not provide a clear explanation of how channel structure changes during gating. While a discrepancy between AlphaFold structural predictions and the experimental measurements is noted, it remains unclear whether this mismatch arises from limitations of the model or from the experimental approach. No further structural analysis is presented to resolve this issue or to clarify the conformational states of the protein.
The manuscript successfully demonstrates that Acd can be incorporated at specific positions without abolishing channel function, and it is noteworthy that the reconstituted proteins function as voltage-activated proton channels in liposomes. The authors also report reversible zinc inhibition of the channel, suggesting that zinc induces structural changes in certain channel regions that can be reversed by EDTA chelation. However, this observation is not explored in sufficient depth to yield meaningful mechanistic insight.
Overall, while the study introduces an interesting labeling strategy and provides valuable methodological observations, the analysis appears incomplete. Additional structural interpretation and mechanistic insight are needed.
Major Points
(1) Tryptophan and tyrosine exhibit similar quantum yields, but their extinction coefficients differ substantially. Is this difference accounted for in your FRET analysis? Please clarify whether this would result in a stronger weighting of tryptophan compared to tyrosine.
(2) Is the fluorescence of acridon-2-ylalanine (Acd) pH-dependent? If so, could local pH variations within the channel environment influence the probe's photophysical properties and affect the measurements?
(3) Several constructs (e.g., K125Tag, Y134Tag, I217Tag, and Q233Tag) display two bands on SDS-PAGE rather than a single band. Could this indicate incomplete translation or premature termination at the introduced tag site? Please clarify.
(4) In Figure 5F, the comparison between predicted FRET values and experimentally determined ratio values appears largely uninformative. The discussion on page 9 suggests either an inaccurate structural model or insufficient quantification of protein dynamics. If the underlying cause cannot be distinguished, how do the authors propose to improve the structural model of hHV1 or better describe its conformational dynamics?
(5) Cu²⁺, Ru²⁺, and Ni²⁺ are presented as suitable FRET acceptors for Acd. Would Zn²⁺ also be expected to function as an acceptor in this context? If so, could structural information be derived from zinc binding independently of Trp/Tyr?
(6) The investigated structure is most likely dimeric. Previous studies report that zinc stabilizes interactions between hHV1 monomers more strongly than in the native dimeric state. Could this provide an explanation for the observed zinc-dependent effects? Additionally, do the detergent micelles used in this study predominantly contain monomers or dimers?
(7) hHV1 normally inserts into a phospholipid bilayer, as used in the reconstitution experiments. In contrast, detergent micelles may form monolayers rather than bilayers. Could the authors clarify the nature of the micelles used and discuss whether the protein is expected to adopt the same fold in a monolayer environment as in a bilayer?
eLife Assessment
This study addresses a fundamental question in glycobiology by elucidating how a single-site processive enzyme orchestrates the alternating addition of sugars to synthesize complex polysaccharides such as hyaluronan. The findings are compelling, providing a clear mechanistic framework supported by strong experimental validation. Major strengths include the integration of high-resolution structural data with rigorous biochemical analyses, resulting in a well-supported model of hyaluronan assembly.
Reviewer #1 (Public review):
Summary:
This revised manuscript describes critical intermediate reaction steps of a HA synthase at the molecular level; specifically, they examine the 2nd step, polymerization, adding GlcA to GlcNAc to form the initial disaccharide of the repeating HA structure. Unlike the vast majority of known glycosyltransferases, the viral HAS (a convenient proxy extrapolated to resemble the vertebrate forms) uses a single pocket to catalyze both monosaccharide transfer steps. The authors work illustrates the interactions needed to bind & proof-read the UDP-GlcA using direct and '2nd layer' amino acid residues. This step also allows the HAS to distinguish the two UDP-sugars; this is very important as the enzymes are not known or observed to make homopolymers of only GlcA or GlcNAc, but only make the HA disaccharide repeats GlcNAc-GlcA.
Strengths:
Techniques & analysis; overview of HA synthase mechanisms
Weaknesses:
None
Comments on revisions:
Previous clarity issues in the original submission were all resolved. Again, this is a very well done body of work!!
Reviewer #2 (Public review):
Summary:
The paper by Stephens and co-workers provides important mechanistic insight into how hyaluronan synthase (HAS) coordinates alternating GlcNAc and GlcA incorporation using a single Type-I catalytic centre. Through cryo-EM structures capturing both "proofreading" and fully "inserted" binding poses of UDP-GlcA, combined with detailed biochemical analysis, the authors show how the enzyme selectively recognizes the GlcA carboxylate, stabilizes substrates through conformational gating, and requires a priming GlcNAc for productive turnover.
These findings clarify how one active site can manage two chemically distinct donor sugars while simultaneously coupling catalysis to polymer translocation.
The work also reports a DDM-bound, detergent-inhibited conformation that possibly illuminates features of the acceptor pocket, although this appears to be a purification artefact (it is indeed inhibitory) rather than a relevant biological state.
Overall, the study convincingly establishes a unified catalytic mechanism for Type-I HAS enzymes and represents a significant advance in understanding HA biosynthesis at the molecular level.
Strengths:
There are many strengths.
This is a multi-disciplinary study with very high-quality cryo-EM and enzyme kinetics (backed up with orthogonal methods of product analysis) to justify the conclusions discussed above.
Comments on revisions:
The suggestions made in the initial comments have all been responded to very well.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
This manuscript describes critical intermediate reaction steps of a HA synthase at the molecular level; specifically, it examines the 2nd step, polymerization, adding GlcA to GlcNAc to form the initial disaccharide of the repeating HA structure. Unlike the vast majority of known glycosyltransferases, the viral HAS (a convenient proxy extrapolated to resemble the vertebrate forms) uses a single pocket to catalyze both monosaccharide transfer steps. The authors' work illustrates the interactions needed to bind & proof-read the UDP-GlcA using direct and '2nd layer' amino acid residues. This step also allows the HAS to distinguish the two UDP-sugars; this is very important as the enzymes are not known or observed to make homopolymers of only GlcA or GlcNAc, but only make the HA disaccharide repeats GlcNAc-GlcA.
Strengths:
Overall, the strengths of this paper lie in its techniques & analysis.
The authors make significant leaps forward towards understanding this process using a variety of tools and comparisons of wild-type & mutant enzymes. The work is well presented overall with respect to the text and illustrations (especially the 3D representations), and the robustness of the analyses & statistics is also noteworthy.
Furthermore, the authors make some strides towards creating novel sugar polymers using alternative primers & work with detergent binding to the HAS. The authors tested a wide variety of monosaccharides and several disaccharides for primer activity and observed that GlcA could be added to cellobiose and chitobiose, which are moderately close structural analogs to HA disaccharides. Did the authors also test the readily available HA tetramer (HA4, [GlcA-GlcNAc]2) as a primer in their system? This is a highly recommended experiment; if it works, then this molecule may also be useful for cryo-EM studies of CvHAS as well.
The reviewer requested testing whether an HA tetratsaccharide could also serve as an glycosyl transfer acceptor for HAS. The commerically available HA tetrasaccharide (HA4) is terminated at its non-reducing end by GlcA, therein we proceeded to measure its effect on UDP-GlcNAc turnover kientics. Titration of HA4 failed to elicit any detectable change in UDP-GlcNAc turnover rate, indicating no priming. This is now mentioned in the main text and the data is shown in Fig. S9.
Weaknesses:
In the past, another report describing the failed attempt of elongating short primers (HA4 & chitin oligosaccharides larger than the cello- or chitobiose that have activity in this report) with a vertebrate HAS, XlHAS1, an enzyme that seems to behave like the CvHAS ( https://pubmed.ncbi.nlm.nih.gov/10473619/); this work should probably be cited and briefly discussed. It may be that the longer primers in the 1999 paper and/or the different construct or isolation specifics (detergent extract vs crude) were not conducive to the extension reaction, as the authors extracted recombinant enzyme.
We apologize for the oversight. This reference is now cited (ref. 18) together with the description of the failed elongation of HA4 by CvHAS.
There are a few areas that should be addressed for clarity and correctness, especially defining the class of HAS studied here (Class I-NR) as the results may (Class I-R) or may not (Class II) align (see comment (a) below), but overall, a very nicely done body of work that will significantly enhance understanding in the field.
Done as requested
Reviewer #2 (Public review):
Summary:
The paper by Stephens and co-workers provides important mechanistic insight into how hyaluronan synthase (HAS) coordinates alternating GlcNAc and GlcA incorporation using a single Type-I catalytic centre. Through cryo-EM structures capturing both "proofreading" and fully "inserted" binding poses of UDP-GlcA, combined with detailed biochemical analysis, the authors show how the enzyme selectively recognizes the GlcA carboxylate, stabilizes substrates through conformational gating, and requires a priming GlcNAc for productive turnover.
These findings clarify how one active site can manage two chemically distinct donor sugars while simultaneously coupling catalysis to polymer translocation.
The work also reports a DDM-bound, detergent-inhibited conformation that possibly illuminates features of the acceptor pocket, although this appears to be a purification artefact (it is indeed inhibitory) rather than a relevant biological state.
Overall, the study convincingly establishes a unified catalytic mechanism for Type-I HAS enzymes and represents a significant advance in understanding HA biosynthesis at the molecular level.
Strengths:
There are many strengths.
This is a multi-disciplinary study with very high-quality cryo-EM and enzyme kinetics (backed up with orthogonal methods of product analysis) to justify the conclusions discussed above.
Weaknesses:
There are few weaknesses.
The abstract and introduction assume a lot of detailed prior knowledge about hyaluronan synthases, and in doing so, risk lessening the readership pool.
A lot of discussion focuses on detergents (whose presence is totally inhibitory) and transfer to non-biological acceptors (at high concentrations). This risks weakening the manuscript.
The abstract and parts of the introduction have been revised to address the reviewer’s concerns.
Reviewer #1 (Recommendations for the authors):
(1) As noted above, please state in title, abstract & introduction that this work is focused on a "Class I-NR HAS" (as described in Ref. #4), and NOT all HAS families...this is truly essential to note as someone working with the Pasteurella HAS version (Class II) would be totally misled & at this point, no one knows the Streptococcus HAS (Class-IR) mechanistic details which could be different due to its inverse molecular directionality of elongation compared to the CvHAS Class I-NR enzyme.
Done as requested.
(2) Page 6 - for the usefulness of the HAS mutants as being folded correctly, it was stated these mutants are suitable since they all 'purify' similarly...the use of the more proper term should probably be 'chromatograph', similarly suggesting similar hydrodynamic radii without massive folding issues.
This has been revised to state that they all exhibited comparable size exclusion chromatography profiles.
“All mutants share similar size exclusion chromatography profiles with the WT enzyme, suggesting that the substitutions do not cause a folding defect (Fig. S3).”
(3) Page 7 - please check these sentences (& rest of paragraph?) as the meaning is not clear. "First, UDP-GlcNAc was titrated in the presence of excess UDP-GlcA, resulting in a response similar to the acceptor-free condition (Fig. 2C). However, the maximum reaction velocity at 20 mM UDP-GlcNAc was approximately 25% lower than that measured in the presence of UDP-GlcNAc only (Fig. 2C)."
The paragraph has been revised to avoid confusion.
(4) In Methods, please use an italicized 'g' for the centrifugation steps globally.
Changed as requested
(5) Please note the source/vendor for the HA standards on gels.
Done
(6) Page 35 - TLC section.
(a) 'n-butanol' (with italic n) is the most widespread chemical name (not butan-1-ol).
Done
(b) Also, for all of the TLC images, the origin and the solvent front should be marked.
Changed as suggested.
Reviewer #2 (Recommendations for the authors):
A number of minor issues should be addressed.
(1) Abstract
Two comments on the Abstract, which I found surprisingly weak given the quality of the work, and lacking a key detail.
A major conceptual contribution of this work is the demonstration of how a single Type-I catalytic centre discriminates, positions, and transfers two chemically distinct substrates in an alternating pattern. This distinguishes HAS from dual-active-site (Type-II) glycosyltransferases and is important for understanding HA polymerization.
However, this central point is not clearly articulated in the abstract. I suggest explicitly stating that HAS performs both GlcNAc and GlcA transfer reactions within a single catalytic site, and that the proofreading/inserted poses illuminate how this multifunctionality is achieved.
The abstract currently ends with the observation of a DDM-bound, detergent-inhibited state. While this is interesting, it absolutely does not represent the central conceptual advance of the study and gives the abstract an artefactual ending.
I strongly recommend revising the final sentences to emphasize the broader mechanistic insight and not an "artefact" (indeed, the enzyme is inactive in the presence of this detergent; it is thus a very unusual way to conclude an abstract).
That is, finish with the wider implications of how HAS coordinates alternating substrate use, proofreading, and polymer translocation. Ending on the main mechanistic or biological significance would make the abstract considerably stronger and more aligned with the main message of the paper.
The abstract has been revised thoroughly to reflect the important insights gained on CvHAS’ catalytic function and HA biogenesis in general.
(2) Introduction
The distinction between single active-centre enzymes, which transfer both sugars alternately, and twin catalytic domain enzymes that each perform one addition is surely central to the whole paper. But it is not discussed. Surely this has to be covered. There is a lot of work in this space, including, but not limited to:
https://doi.org/10.1093/glycob/cwg085
https://doi.org/10.1093/glycob/10.9.883
https://doi.org/10.1093/glycob/cwad075 (includes this author team)
Originally back to https://doi.org/10.1021/bi990270y
If the authors instead assume such a level of knowledge for the reader, then surely they are writing for a specialist audience, not consistent with the wider readership ambitions of eLife?
The Introduction has been revised as suggested by the reviewer, providing necessary background to frame our description of the Chlorella virus HAS. We made a deliberate effort to put new insights into a broader context.
(3) Results and Discussion
DDM "was observed for >50% of the analysed particles". I struggled with this. I couldn't understand how the authors selected particles that did or did not contain DDM. The main body text states: "To our surprise, careful sorting of the UDP-GlcA supplemented cryo EM dataset revealed a CvHAS subpopulation that was not bound to the substrate, but, instead, a DDM molecule near the active site (Fig 3A and S7). This was observed for >50% of the analyzed particles."
That reads like there is one sample with two populations. But the figures and the methods section suggest differently: they suggest two samples with different data-collection regimes. That does not match the main text. Could this be clarified?
Yes, that wasn’t explained well. We clarified the text to stress that the DDM-bound sample came from a dataset that was intended to resolve an UDP-GlcA-bound state, but instead revealed the inhibition by DDM.
Also in this space, in the modern world, "nominal magnification" has no real meaning, and calibrated pixel size would be more appropriate. Can this be given, please?
The relevant Methods section now states: “imaging of … was performed at a calibrated pixel size of 0.652 Å”.
The discovery of DDM in the active site is surprising. But it is an inhibitory artefact. Is this section pushed a little too hard? Also, "The coordination of DDM's maltoside moiety, an αlinked glucose disaccharide, is consistent with priming by cellobiose and chitobiose." I'm not sure why an α-linked maltose is consistent with the binding of a β-linked cellobiose. That makes no sense. There will be no other enzymes where starch and cellulose oligos are mutually accepted. Consider rewriting.
We like to stress the DDM coordination because it could lead to the development of compounds that can really function as inhibitors, either for HAS or other related enzymes. In the observed DDM binding pose, the alpha-linkage is not recognized. Instead, the reducing end glucosyl unit stacks against Trp342 while the non-reducing unit extends into the catalytic pocket. Hence, a similar binding pose is conceivable for cellobiose and potentially also for chitobiose. The relevant section has been reworded.
eLife Assessment
This important study introduces an approach to discovering antibiotic resistance determinants by leveraging diverse susceptibility profiles among related mycobacterial species, with particular relevance to high-level resistance against natural product-derived antibiotics. The research provides convincing evidence for the role of ADP-ribosylation enzymes in rifamycin resistance among mycobacteria, whilst also demonstrating that antibiotic susceptibility is not correlated with growth rate or intracellular compound concentration. The revision is substantively improved, but some broader claims still require additional experimental support. This work lays a significant foundation for understanding the complexity of antibiotic resistance mechanisms in mycobacteria and opens new avenues for future antimicrobial research.
Reviewer #1 (Public review):
This work analyzes innate resistance to drugs in mycobacteria by comparing minimum inhibitory concentrations (MICs) across a diverse panel of mycobacterial species. The results show that MICs are poorly correlated with growth rate while phylogeny associated with horizontal gene transfer underlies the observed differences in MIC, an important demonstration. A further investigation into the driver for the vast differences in susceptibility profiles shows that for three drugs the MIC is not correlated with intrabacterial drug concentrations where intrabacterial drug concentration is comprised of cytosolic and cell wall associated drug. This is a striking observation. The authors delve into the mechanisms that drive resistance to rifamycins and confirm that resistance is driven by ADP-ribosyltransferases of which two variant groups exist, one of which is kinetically faster and apparently is superior at modifying more hydrophobic rifamycins. The relative role of the two ADP-ribosyltransferases in conferring resistance especially in the species with both orthologs is not fully understood since the modified drug can possibly be further modified and transcriptional downregulation experiments performed in this work do not provide genetic evidence of perturbation of mRNA levels of the respective open reading frames.
Comments on revisions:
Demonstration of the level of transcriptional downregulation of the two Arr orthologs would have been a nice demonstration of (1) the utility of CRISPRi in other mycobacteria, (2) that the difference in rifabutin susceptibility during knockdown of Arr-1 vs Arr-X can fully be ascribed to the role of Arr-X in modifying the drug.
Reviewer #3 (Public review):
This manuscript presents a macroevolutionary approach to identification of novel high-level antibiotic resistance determinants that takes advantage of the natural genetic diversity within a genus (mycobacteria, in this case) by comparing antibiotic resistance profiles across related bacterial species and then using computational, molecular, and cellular approaches to identify and characterize the distinguishing mechanisms of resistance. The approach is contrasted with "microevolutionary" approaches based on comparing resistant and susceptible strains of the same species and approaches based on ecological sampling that may not include clinically relevant pathogens or related species. The potential for new discoveries with the macroevolution-inspired approach is evident in the diversity of drug susceptibility profiles revealed amongst the selected mycobacterial species and the identification and characterization of a new group of rifamycin-modifying ADP-ribosyltransferase (Arr) orthologs of previously described mycobacterial Arr enzymes. Additional findings that intra-bacterial antibiotic accumulation does not always predict potency within this genus, that M. marinum is a better proxy for M. tuberculosis drug susceptibility than the commonly used saprophyte M. smegmatis, and that susceptibility to semi-synthetic antibiotic classes is generally less variable than susceptibility to antibiotics more directly derived from natural products strengthen the claim that the macroevolutionary lens is valuable for elucidating general principles of susceptibility within a genus.
There are some limitations to the work. The argument for the novelty of the approach could be better articulated. While the opportunities for new discoveries presented by identification of discrepant susceptibility results between related species is evident, it is less clear how the macroevolutionary approach is further leveraged for the discovery of truly novel resistance mechanisms. The example of the discovery of Arr-X enzymes presented here relied upon foundational knowledge of previously characterized Arr orthologs. There is less clarity about what the pipeline would look like for discovery of previously unknown determinants when one is agnostic to putative mechanisms. From the point at which interspecies differences in susceptibility are noted, does the framework still remain distinct from other discovery frameworks and approaches?
While the experimentation and analyses performed are generally well designed and rigorous, there are a few instances in which broad claims are based on inferences from sample sets or data sets that are, at present, too limited to provide robust support. For example, the claim that rifampicin modification, and precisely ADP-ribosylation, is the dominant mechanism of resistance to rifampicin in mycobacteria is still a bit premature or at least an over-generalization, as other enzymatic modification mechanisms and other mechanisms such as helR-mediated dissociation of rifampicin-stalled RNA polymerases, efflux, etc were not examined. CRISPR interference was used in a demonstrative example to support this assertion, but would need to be applied more systematically to be more conclusive. The general claim that intra-bacterial antibiotic accumulation does not predict potency in mycobacteria may be another over-generalization based on the limited set of drugs and species studied.
Comments on revisions:
Discussion, lines 321-323: "We found that resistance to these antibiotics in mycobacteria do not correlate with by uptake/efflux mechanisms in the species tested..." is an over-generalization and conflicts with the following statement on lines 199-201: "for BDQ we could observe some correlation between antibiotic potency and [BDQ]IB which could be indicative of efflux playing a role in antibiotic efficacy. Given that the current statement in the Discussion only applies to 2 of 3 drugs tested, a more specific or nuanced interpretation seems warranted.
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
This work shows that resistance profiles to a variety of drugs are variable between different mycobacterial species and are not correlated with growth rate or intrabacterial compound concentration (at least for linezolid, bedaquiline, and Rifampicin). Note that intrabacterial compound concentration does not distinguish between cytosolic and periplasmic/cell wall-associated drugs. The susceptibility profiles for a wide range of mycobacteria tested under the same conditions against 15 commonly used antimycobacterial drugs provide the first recorded cross-species comparison which will be a valuable resource for the scientific community. To understand the reasons for the high Rifampicin resistance seen in many mycobacteria, the authors confirm the presence of the arr gene known to encode a Rif ribosyltransferase involved in Rif resistance in M. smegmatis in the resistant mycobacteria after confirming the absence of on-target mutations in the RpoB RRDR. Metabolomic analyses confirm the presence of ribosylated Rif in some of the naturally resistant mycobacteria which may not be entirely surprising but an important confirmation. Presumably M. branderi is highly resistant despite lacking the arr homolog due to the rpoB S45N mutation. M. flavescens has an MIC similar to that of M. smegmatis, despite having both Arr-1 and Arr-X. Various Arr-1 and Arr-X proteins are expressed and characterized for catalytic activity which shows that Arr-X is a faster enzyme,, especially with respect to more hydrophobic rifamycins. M. flavescens has similar MIC values to Rifapentine and Rifabutin to M. smegmatis. Thus, the Arr-1 versus Arr-X comparison does not provide a complete explanation for the underlying reasons driving natural Rif resistance in mycobacteria. Downregulation of Arr-X expression in M. conceptionense confers increased sensitivity to Rifabutin confirming its role as a rifamycin-inactivating enzyme.
Overall, the comparison of cross-species susceptibility profiles is novel; the demonstration that MIC is not correlated with intracellular drug concentration is important but not sufficiently interrogated, the demonstration that Arr-X is also a Rif ADP-ribosyltransferase is a good confirmation and shows that it is more efficient than Arr-1 on hydrophobic rifamycins is interesting but maybe not entirely surprising. The manuscript seems to have two parts that are related, but the rifamycin modification aspect of the work is not strongly linked to the first part since it interrogates the modification of one drug but not the common cause of natural resistance for other drugs.
Reviewer #2 (Public review):
Summary:
The authors use a variety of methods to investigate the mechanisms of innate drug resistance in mycobacteria. They end up focusing on two primary determinants - drug accumulation, which correlates rather poorly with resistance for many species, and, for the rifamycins, ADP-ribosyltransferases. The latter enzymes do appear to account for a good deal of resistance, though it is difficult to extrapolate quantitatively what their relative contributions are.
Overall, they make excellent use of biochemical methods to support their conclusions. Though they set out to draw very broad lessons, much of the focus ends up being on rifamycins. This is still a very interesting set of conclusions.
Strengths:
(1) A very interesting approach and set of questions.
(2) Outstanding technical approaches to measuring intracellular drug concentrations and chemical modification of rifamycins.
(3) Excellent characterization of variant rifamycin ADP-ribosyltransferases
Weaknesses:
(1) Figure 3c/d: These panels show the same experiment done twice, yet they display substantially different results in certain cases. For instance, M. smegmatis appears to show an order of magnitude lower RIF accumulation in panel d compared to M. flavescens, despite them displaying equal accumulation in panel c. The authors should provide justification for this variation, particularly as quantitative intra-species comparisons are central to the conclusions of this figure.
The data in panels 3c and 3d are from different sets of experiments. The reviewer is correct with regards to M. smegmatis. The data indeed is ~ 1 order of magnitude different. However, the data for other species is very similar. The reviewer may also have noticed that the error bars are also larger in 3d, compared to 3c, indicating a greater variation between independent experiments use in 3d. We do not have a good explanation for this, other than the experiments shown in 3d were associated with greater biological variability.
(2) There are several technical concerns with Figure 3 that affect how to interpret the work. According to the methods, the authors did not appear to normalize to an internal standard, only to an external antibiotic standard (which may account for some of the technical variation alluded to above).
We agree that using a labeled drug as an internal standard (IS) would be ideal. However, the experiment initially followed an untargeted metabolomics approach, which later shifted to relative drug quantification. At that stage, normalizing with IS was impractical because proper implementation would require multiple IS across the chromatographic range. Therefore, we opted for total ion current (TIC) normalization, which accounts for variability in overall metabolite abundance—even though the experimental setup was already adjusted for each bacterial species’ growth rate. Additionally, we prepared external standard curves for each drug to enable quantification, and the amount of drug added to each plate was considered when reporting these values.
Second, the authors used different concentrations of drug for each species to try to match the species' MICs. I appreciate the authors' thinking on this, but I think for an uptake experiment it would be more appropriate to treat with the same concentration of drug since uptake is likely saturable at higher drug concentrations. In the current setup, for the species with higher MIC, they have to be able to uptake substantially more antibiotics than the species with low MIC in order to end up with the same normalized uptake value in Figure 3d. It would be helpful to repeat this experiment with a single drug concentration in the media for all species and test whether that gives the same results seen here.
We respectfully disagree with the reviewer. Experiments such as the one proposed by the review work well when MIC values are a few fold apart, for strains of the same species, but have not been tested when MIC values are 100-1000-fold apart, with different species. Furthermore, what would be the interpretation of compound uptake at 1000-fold the MIC for one species and MIC level for another? By using antibiotic concentrations at the respective MIC for each species we are at least under conditions where we know the biological effect of the antibiotic across species is the same, based on its potency.
(3) Figure 4f: This panel seems to argue against the idea that the efficacy of RIF ribosylation is what's driving drug susceptibility. M. flavescens is similarly resistant to RIF as M. smegmatis, yet M. flavescens has dramatically lower riboslyation of RIF. This is perhaps not surprising, as the authors appropriately highlight the number of different rif-modifying enzymes that have been identified that likely also contribute to drug resistance. However, I do think this means that the authors can't make the claim that the resistance they observe is caused by rifamycin modification, so those claims in the text and figure legend should be altered unless the authors can provide further evidence to support them. This experiment also has results that are inconsistent with what appears to be an identical experiment performed in Supplemental Figure 5b. The authors should provide context for why these results differ.
In regard to enzyme efficiency, the apparent rate of all Arr-1 is relatively similar in converting RIF into ADP-Ribosyl-Rif between species. However, Arr-X is much more efficient when compared to Arr-1 in both M. flavescents and M. conceptionense. This is indicated by the apparent rate measured and displayed on figure 5c.
Proteomics data shows that there is upregulation of Arr-1 and Arr-X upon rifampicin treatment in M. flavescens and M. conceptionense. However, the same experiment was not performed in Arr-1 KD. Therefore, we can’t verify through this approach if the activity observed in vivo directly correlates with a higher expression of Arr-X alone. Of note, likely both enzymes contribute to resistance to rifamycins, as per our results with the Arr-X KD and sensitization of M. conceptionense to RIF.
Author response image 1.
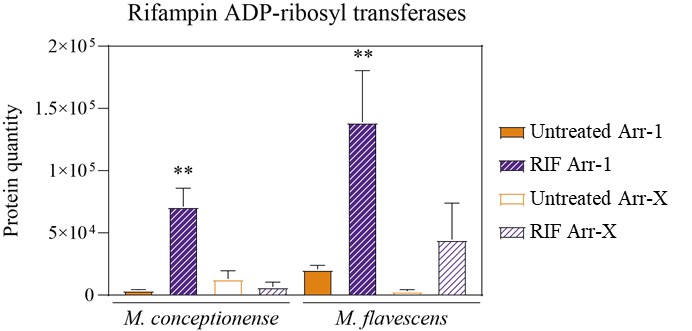
It is also worth mentioning that there are other enzymes in the pathway of RIF ribosylation and their efficiency is unknown (Author response image 2). Therefore ADP-Ribosyl-RIF It is not an “end-metabolite” and maybe not the sole determinant of RIF resistance via ADP-ribosylation. Downstream enzymes can also account for the difference observed between M. flavescens and M. smegmatis.
Author response image 2.
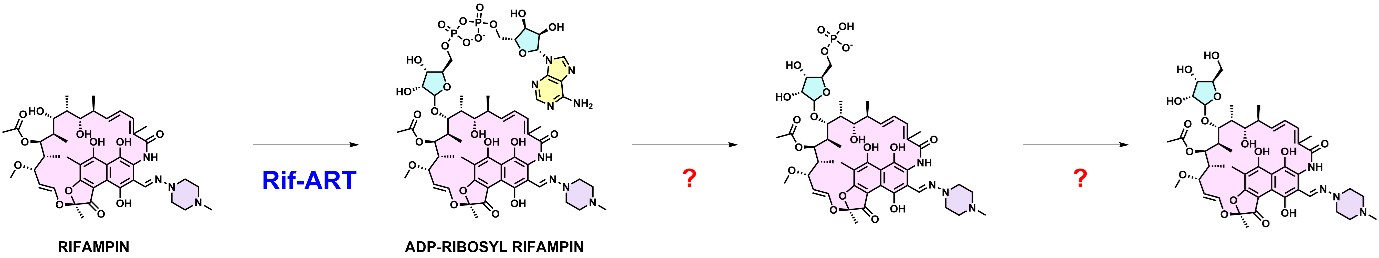
It is correct that the Rifampicin MIC for M. flavescens is the same as M. smegmatis.
(4) Fig 4f/5c: M. flavescens has both Arr-1 and Arr-X, yet it appears to not have ribosylated RIF. This result seems to undermine the authors' reliance on the enzyme assay shown in Fig 5c - in that assay, M. flavescens Arr-X is very capable of modifying rifampicin, yet that doesn't appear to translate to the in vivo setting. This is of importance because the authors use this enzyme assay to argue that Arr-X is a fundamentally more powerful RIF resistance mechanism than Arr-1 and that it has specificity for rifabutin. However, the result in Figure 4f would argue that the enzyme assay results cannot be directly translated to in vivo contexts. For the authors to claim that Arr-X is most potent at modifying rifabutin, they could test their CRISPRi knockdowns of Arr-X and Arr-1 under treatment with each of the rifamycins they use in the enzyme assay. The authors mentioned that they didn't do this because all the strains are resistant to those compounds; however, if Arr-X is important for drug resistance, it would be reasonable to expect to see sensitization of the bacteria to those compounds upon knockdown.
The reviewer is reading Fig. 4f incorrectly, probably because it is plotted in a linear scale instead of logarithmic scale. Ribosylated Rif is present in M. flavescens, just at lower levels than M. conceptionense and M. smegmatis. In species where there is no Arr-1 or Arr-3, ribosylated RIF is not detected at all (e.g. M. tuberculosis), i.e., concentration is zero. Therefore, any detection of ribosylated RIF can be considered significant. In addition, as mentioned before, ADP-ribosylation of RIF is not the final product of the reaction and further studies need to be undertaken to understand subsequent reactions.
(5) Figure 5d: The authors use this CRISRPi experiment to claim that ArrX from M. conceptionanse is more potent at inactivating rifabutin than Arr-1. This claim depends on there being equal degrees of knockdown of Arr-1 and Arr-X, so the authors should validate the degree of knockdown they get. This is particularly important because, to my knowledge, nobody has used this system in M. conceptionanse before.
We agree with the reviewer that a qPCR should have been performed to define the extent of interference in the strain. generated Unfortunately, at this time a qPCR was not performed in the strains tested to confirm the extent of down regulation. Although it is the best practice to validate the strain KD, there is no indication that the effect observed is due to unspecific downregulation. The genetic environment in which Arr-X is positioned is different from Arr-1 and the targeting oligonucleotides are specific and would not promiscuously bind to Arr-1. Said that, this is indeed a fault in our setup.
(6) The authors' arguments about Arr-X and Arr-1 would be strengthened by showing by LC/MS that Arr-X knockdown in M. conceptionense results in more loss of ribosyl-rifabutin than knockdown of Arr-1.
We agree with the reviewer that performing the LC-MS analysis of the Arr-x knockdown would have strengthened the argument of our paper. Unfortunately, this experiment was not performed.
Reviewer #3 (Public review):
This manuscript presents a macroevolutionary approach to the identification of novel high-level antibiotic resistance determinants that takes advantage of the natural genetic diversity within a genus (mycobacteria, in this case) by comparing antibiotic resistance profiles across related bacterial species and then using computational, molecular, and cellular approaches to identify and characterize the distinguishing mechanisms of resistance. The approach is contrasted with "microevolutionary" approaches based on comparing resistant and susceptible strains of the same species and approaches based on ecological sampling that may not include clinically relevant pathogens or related species. The potential for new discoveries with the macroevolution-inspired approach is evident in the diversity of drug susceptibility profiles revealed amongst the selected mycobacterial species and the identification and characterization of a new group of rifamycin-modifying ADP-ribosyltransferase (Arr) orthologs of previously described mycobacterial Arr enzymes. Additional findings that intra-bacterial antibiotic accumulation does not always predict potency within this genus, that M. marinum is a better proxy for M. tuberculosis drug susceptibility than the commonly used saprophyte M. smegmatis, and that susceptibility to semi-synthetic antibiotic classes is generally less variable than susceptibility to antibiotics more directly derived from natural products strengthen the claim that the macroevolutionary lens is valuable for elucidating general principles of susceptibility within a genus.
There are some limitations to the work. The argument for the novelty of the approach could be better articulated. While the opportunities for new discoveries presented by the identification of discrepant susceptibility results between related species are evident, it is less clear how the macroevolutionary approach is further leveraged for the discovery of truly novel resistance determinants. The example of the discovery of Arr-X enzymes presented here relied upon foundational knowledge of previously characterized Arr orthologs. There is little clarity on what the pipeline for identifying more novel resistance determinants would look like. In other words, what does the macroevolutionary perspective contribute to discovery from the point of finding interspecies differences in susceptibility? Does the framework still remain distinct from other discovery frameworks and approaches? If so, how?
Thanks for pointing this out, as this is a critical feature of our study and method. Our approach relies on inter-species comparative genomics and phenotypes, and therefore, it is distinct from inter-strains comparison. This difference is dramatic, and it becomes clearer when we are comparing the core genome of M. tuberculosis (one species) 92% with the core genome of the genus, circa of 1%. While we focus on rifamycin in this manuscript, future manuscripts will investigate many of the other dozens of “inconsistencies” observed between the genetic makeup of different mycobacterial species and there actual performance in the presence of different antibiotics.
While the experimentation and analyses performed appear well-designed and rigorous, there are a few instances in which broad claims are based on inferences from sample sets or data sets that are too limited to provide robust support. For example, the claim that rifampicin modification, and precisely ADP-ribosylation, is the dominant mechanism of resistance to rifampicin in mycobacteria may be a bit premature or an over-generalization, as other enzymatic modification mechanisms and other mechanisms such as helR-mediated dissociation of rifampicin-stalled RNA polymerases, efflux, etc were not examined nor were CRISPRi knockdown experiments conducted beyond an experiment to tease out the role of Arr-X and Arr-1 in one strain. The general claim that intra-bacterial antibiotic accumulation does not predict potency in mycobacteria may be another over-generalization based on the limited number of drugs and species studied, but perhaps the intended assertion was that antibiotic accumulation ALONE does not predict potency.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Major comments
(1) The metabolomics is done using mycobacteria grown on filters. Initially, mycobacterial cells are grown on the filters for 5 doublings before being transferred to drug-containing (or free) agar for one doubling. Is this based on calculated doubling time in liquid culture or a true determination of the fact that the biomass increases to what would amount to 5 doublings?
The doubling time used is the one determined in liquid media. Although it is possible that the growth kinetics in solid media is slightly different from liquid (±10%), this experimental design is well established for M. tuberculosis (since Proc Natl Acad Sci U S A. 2010 May 25;107(21):9819-24.) and M. smegmatis (unpublished). Therefore, we used the growth rate as a proxy for having the same biomass of cells for each species tested. A maximum difference of 10% was observed between M. tuberculosis growth in liquid and in solid media, however, cells grow exponentially for much longer in filters. This makes filter-based experiments more reliable, as few growth phase-derived differences are present.
(2) The demonstration that intrabacterial drug concentrations vary between mycobacterial species in a manner not related to MIC for at least LZD and RIF, is an important finding. However, intrabacterial does not mean cytoplasmic since a considerable fraction could be present in the periplasmic/cell wall layers. Ideally, this would need to be determined but would of course be a massive undertaking since the method needs validation & optimization for each mycobacterial species. Nevertheless, this has to be mentioned. In addition, three drugs are limiting. Measuring additional drug concentrations in these 5 mycobacteria would at least establish some confirmation about the extent of this lack of correlation. Thus, could the authors measure concentrations of additional drugs with intracellular targets?
Testing additional drugs can be beneficial and would be an expansion of our paper, which will definitely be on future plans for further studies focusing on other antibiotics described here. It would also provide new insights into other possible mechanisms of resistance in mycobacterial species. However, in this study we aimed to first determine the antibiotic response profile in different mycobacterial species, and once we identified interesting resistance phenotypes that could not be readily explained by known mechanisms of resistance, we narrowed it down to certain drugs and species that would potentially provide insights into new mechanisms of antibiotic resistance. Finally, exploring drug concentration across multiple bacterial compartments is a dauting task and it has not been done extensively with any species, not to mention with multiple species, many of which are still lacking any study of their actual cell envelope.
(3) CRISPRi was used to reduce transcription in M. conceptionense. What was the level of gene downregulation?
As mentioned previously, a setback from our setup is that the level of KD was not measured at this instance.
Minor comments:
(1) The introduction mentions the fast and slow-growing mycobacteria which are classified based on the time that it takes to observe colonies on solid agar. However, in liquid medium, there is less correlation between the reported growth on agar and doubling time in liquid (Figure 1b, Figure 2d). This could be mentioned in the results section. In Figure 2d, the filled circles represent fast-growers but this does not hold well for liquid culture and it might make more sense to not distinguish between fast- and slow-growers in these graphs. A small complication would also be the fact that the doubling time represents growth in a liquid medium with Tyloxapol as a detergent whereas the MIC and metabolomics are done on solid agar with no detergent. The metabolomics is done after a doubling but for those where agar growth and liquid growth have large discrepancies in growth rate, there could be some differences.
Apologies for this misunderstanding. Fast- and slow-growth phenotypes are determined in Lowenstein-Jensen (LJ) agar, not in 7H10 agar (used in our study and most studies of mycobacteria). Furthermore, this is a qualitative definition, not a quantitative one. Therefore, our measurements do not need to correlate with fast- and slow-growth phenotypes, unless we had used that one specific medium. Furthermore, in liquid medium, we determined growth rate directly, which is never done with LJ medium.
In addition to adding the same amount of cells to each filter, we also perform TIC normalization, which should account for how rich the samples were – and therefore how much material we had. Therefore, we do not observe discrepancies due to differences in growth rate and the presence/absence of detergent in the media.
It is also worth mentioning that this experimental set up has been well established in many M. tuberculosis labs that study metabolism. Importantly, the use of detergent drastically affects mass spectrometry, and therefore cannot be used.
(2) Figure 1g in the text should be Figure 1f.
Apologies, it has been fixed.
(3) Figure S1 would be ideal to have in (supplementary) table format.
This data is now being provided in a table format.
(4) Table S1 - ethambutol misspelt.
Spelling has been corrected.
(5) MIC for species such as M. abscessus could depend on medium (7H9-based medium can give different MIC values than CAMH).
Indeed, different media can significantly change MIC values, and this is true for many bacterial species, if not all. For this study we used only species that could be grown in 7H9 broth containing 10 % ADC, 0.05% glycerol 0.05% tyloxapol and 7H10 plates containing 10% OADC and 0.05% glycerol. MIC<sub>99</sub> was determined in the latter as we found more efficient and robust to do our tests it in solid media. The goal of our experiment was not to the determined the “true” MIC for the antibiotics tested, as this value does not exist. It was to find lack of correlations between relative values and the presence of genes that can account for it.
(6) The statement "the experiment was performed at a concentration of antibiotic equal to its MIC" initially seems confusing. It was not equal to the MIC but performed at 6-fold the respective MIC of the species in question. Maybe re-phrasing this would help.
Apologies for this oversight. It has been corrected.
(7) Note that some mutations outside the RRDR (eg. V170F and I491F) can also cause Rif resistance.
Author response image 3.
A Rainbow diagram of RpoB X-Ray structure coloured according to sequence conservation. Dark purple indicates high conservation, whereas dark orange indicates low conservation. RIF (showed in magenta) is bound to RpoB. Zoomed view displays that the RIF-binding pocket is considerably conserved. B RpoB protein sequence has an 81bp region called Rifampicin Resistance Determining Region (RRDR) that is known to be important for RIF binding and is where most mutations occur in drug-resistant TB. Sequence alignment displays that the RRDR region is conserved with the exception of M. branderi, which has an Asn instead of a Ser residue in position 456 (numbering is related to the M. tuberculosis sequence), highlighted in bold.
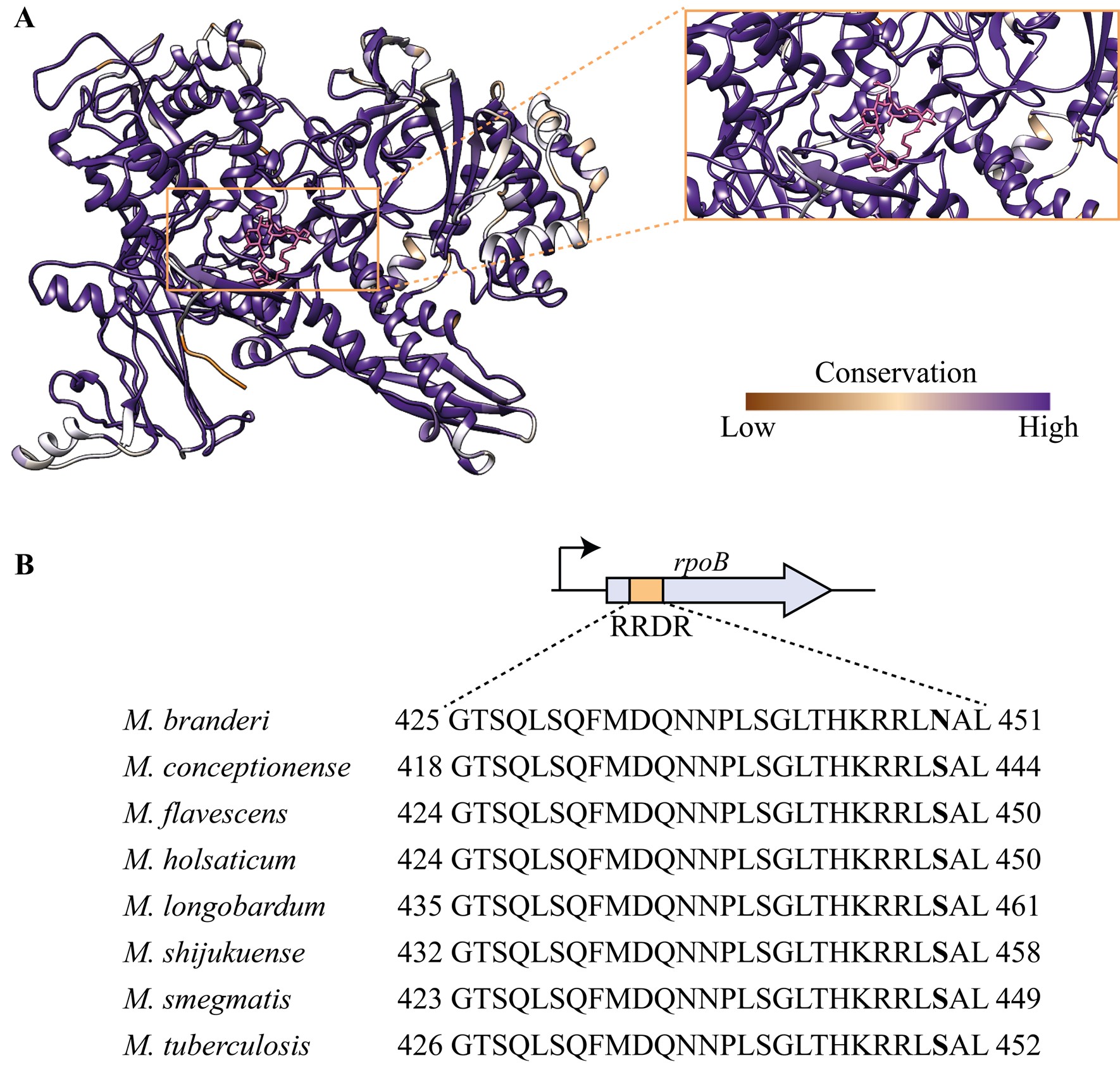
Attached we have a structural alignment of RpoB of the species highlighted on this paper. Although there is variability within the sequences, which is also displayed in Author response image 3 with the conservation analysis, the residues that have been implicated with resistance (including V170 and I491) are conserved. Alignment sent on .fasta file that can be opened in jalview.
(8) Discuss how the RpoB S450N mutation in M. branderi confers the observed level of resistance.
That’s a great point, thank you. Now it reads as:
“The rifampicin (RIF) binding pocket is generally conserved, but Mycobacterium branderi has an S450N mutation in the RRDR region. While this specific mutation hasn't been found in clinical isolates, it's located at the binding site and may confer resistance (273). Although both serine (S) and asparagine (N) have similar side chains, related mutations like S450Q have been linked to resistance (156). Thus, M. branderi may be RIF-resistant due to this mutation. In contrast, M. conceptionense, M. flavescens, and M. smegmatis show no target sequence differences that explain their resistance”
(9) The statement that the three tested NTM are sensitive to rifabutin ("resistant to all rifamycins except for rifabutin") needs to be interpreted considering what sensitivity means. The MIC is still high (1.6-3.1 ug/mL) when compared to that of Mtb. The 2-fold differences in MIC between M. smegmatis and M. conceptionense do not really prove or disprove the role of Arr-X in rifabutin resistance.
We fixed the sentence to be more careful with the language on the text. We agree, but it is worth mentioning that generally with bacteria there is a regulation by the CLSI. Each bacterial species has a range that is considered sensitive or resistant, but these are not available for the species used in this study. In general, bacteria with MIC values above 8 µg/mL are considered resistant to rifampin (J Antibiot 2014 67:625).
(10) Figure 1d: It's hard to quantify the sensitivity of the plates. Can this be done by MIC? Was only rifabutin tested or also rifampicin?
The initial experiments described on the paper were all performed using Rifampicin only. Then, the MIC for the remaining rifamycins was determined for M. smegmatis, M. flavescens and M. conceptionense, and can be perused on “Supplementary table 4”. Figure 5d is to illustrate the effect of the KD in M. conceptionense sensitivity to rifabutin.
(11) Is there data to show the ADP-ribosylation of rifabutin in M. conceptionense and the CRISPRi strains?
Unfortunately, we did not perform LC-MS analysis on M. conceptionense CRISPRi strains exposed to rifabutin to measure potential ADP-ribosylation.
Reviewer #2 (Recommendations for the authors):
(1) It would be useful if the authors would complete Figure 1A by determining growth rates for the remaining 18 strains that they currently omitted.
These growth rates were obtained using roller bottles and in at least 3 independent experiments, unfortunately the throughput is far ideal. The goal of the experiment was to highlight difference in growth rate, beyond fast- and slow-growth, which we did. Adding the remaining values would not change this conclusion. Growth rate variation in 7H9 is significant and the point is made in our figure.
(2) The authors should justify their choice of species used in Figures 3-4. It would be useful to know, for instance, if the authors chose these species in an unbiased fashion, or if they were chosen because the authors had already determined that they possess rifamycin-modifying enzymes of interest. In that case, they wouldn't necessarily be a representative sample to use for the correlation analysis of antibiotic uptake and potency in Figure 3.
They were chosen because of their resistance profile for BDQ, LZD and RIF. This has been addressed in the text, which now reads “Given the antibiotic response profiles observed, we selected BDQ, LZD and RIF to explore the molecular causes of these dramatic changes in antibiotic potency observed across the Mycobacterium genus.”
(3) Figure 4b: The data in this panel appear inconsistent - for instance, M. houstonense appears to grow at 10X Mtb MIC, but fails to grow at 1X Mtb MIC. Repeating this experiment would better establish the validity of the authors' claims about the relative susceptibility of these strains to RIF.
The figures got rotated when exported from illustrator. Corrected figure is uploaded, and original plate photos are also uploaded for clarity.
(4) Figure 4e: Does Arr-X get upregulated in these proteomic datasets? The authors' argument that proteomic upregulation correlates with important drug resistance genes would imply that it might be, so that would be useful information to provide.
Arr-X is slightly upregulated, but not statistically significant – this could be due to the native expression of Arr-1. Data is displayed in a previous answer.
(5) I wasn't able to find the supplementary tables that the authors allude to - not sure if that was a file mixup, but those tables would be useful for interpreting the manuscript.
We are sorry that you couldn’t access the table. It must be a file corruption issues, as the other reviewers were able to. We will make sure that all tables are available and accessible.
(6) For LC/MS, the authors use peak height instead of peak area, which they argue correlates better with the amount of drug in cells because of the poor peak shape they observed for linezolid. This is not standard practice, so the authors should provide evidence to support this claim by running an LC/MS standard curve, then showing the correlation between peak height and amount of compound added as well as the correlation between peak area and compound.
Thank you for pointing that out, accuracy calculated and displayed. Both peak area and height can be used, but indeed area is standard practice.
(7) The authors should provide methods information about the LC column and the gradient settings used for LC-MS, as well as the settings of the MS.
The full method has been added to the paper.
Reviewer #3 (Recommendations for the authors):
I have only minor comments aside from the information in the Public Review:
(1) Results, section on Intra-bacterial antibiotic accumulation, line 8: "experiment was performed at a concentration of antibiotic PROPORTIONAL to its MIC" would be more accurate?
Agreed and adjusted according to Reviewer’s suggestion.
(2) Results, section on A minor role for pre-existing target modification, last sentence: the mere presence of RIF-ribosylating enzymes does not, in and of itself indicate that "RIF modification, and precisely ADP-ribosylation, is the dominant mechanism of resistance to RIF in mycobacteria", as other mechanisms and other forms of modifying enzymes are known to confer rifamycin resistance, with redundancy (e.g., other rifampicin-modifying enzymes, or helR-mediated dissociation of rifampicin-stalled RNA polymerases from DNA). It would be more appropriate to suggest the results presented to this point indicate RIF modification is common among mycobacteria. The evidence from the CRISPRi knockdown of Arrs shown in Fig 5d is the kind of evidence that suggests ribosylation as a dominant mechanism, at least against rifabutin in this particular species.
Absolutely, there are other possible modifying enzymes that could be encoded by these mycobacterial species. There is a possibility that M. flavescens and M. smegmatis encode for a putative helR (attached alignment) but further experiments would need to be carried out to confirm its ability to displace RIF in the RNAP. Interestingly, the presence of both Arr and HelR has been studied in M. abscessus and those mechanisms of resistance are independent from each other (Molecular Cell 2022 82(17):3166-3177.e5).
(3) Discussion, 2nd sentence needs grammatical editing.
Rephrased and it reads “Using our mycobacterial library, we identified for the first time high- and ultra-high-level intrinsic resistance (3) to many of the antibiotics tested. Of note, the resistant phenotype is naturally occurring and not a result of mutations due to exposure to the antibiotic in the clinic – which is the more traditional approach for probing mechanisms of antibiotic resistance. Our observations revealed that resistance profiles are highly variable across the genus and do not follow phylogeny, implicating HGT as the key mechanism for acquisition of resistance determinants and evolution of antibiotic resistance in mycobacteria (42).”
(4) Discussion, page 7, first line: the inclusion of LZD and BDQ in this statement seems at odds with Figure 2c and the statements in the first paragraph of page 5 highlighting these as examples of drugs to which most mycobacteria are susceptible.
Indeed, many of the species are susceptible, however the MIC<sub>99</sub> levels observed have never been reported before, and therefore we found it to be an interesting finding to highlight. From a treatment perspective, knowing which species are sensitive to which drugs is of course the most useful outcome of our study.
(5) The next sentence..."We found that resistance to these antibiotics in mycobacteria cannot be explained by uptake/efflux mechanisms..." is a bit of an over-generalization and conflicts with the evidence presented earlier that efflux could be playing a role in BDQ resistance and the published evidence establishing a clinically significant role for efflux-mediated BDQ resistance in M. tuberculosis, M. avium complex and M. abscessus complex.
We rephrased it to make it more specific to our findings. It reads “We found that resistance to these antibiotics in mycobacteria do not correlate with by uptake/efflux mechanisms in the species tested and it does not correlate with growth rate. Identification of mycobacterial species highly resistant to BDQ and LZD is worrisome as most of this species, if not all, have never been exposed to these drugs.”
(6) Methods, section on In vitro activity assay of Arr enzymes, line 1: reference(s) should be provided for previously reported methods.
Reference now added.
(7) Figure 2d: the low end of the susceptibility range is not well defined.
In this figure the susceptibility is not defined as the lowest area of the graph, but the lower concentrations are indeed harder to be defined. Hopefully supplementary figure 1 and the additional table containing the MIC can be informative to address this comment.
(8) Figures 3c,d: the presentation of the relative antibiotic concentrations could be harmonized between the graphs in 3c and those in 3d to enable a more ready comparison.
We disagree. The goal of these different panels is exactly to illustrate two distinct points. C gives the relative concentration of antibiotic, while D correlates relative concentration with MIC99. The use of log scale in D further clarifies that there is no correlation between intracellular antibiotic concentration and potency (MIC). This information is not present in C.
(9) Figure 4f and Supplementary Figure 5b: it is difficult to understand the limited amount of ribsosyl-RIF in M. flavescens in Fig 4f relative to Supplementary Figure 5b (esp. when considering M. smeg as a common comparator); and, further, to understand the seeming lack of correlation between RIF susceptibility, ribosylation and Arr number and catalytic efficiency for these two strains without considering additional resistance mechanisms.
In reality the difference between figure 4f and Supplementary figure 5b is mainly due to M. smegmatis – that has an apparent lower production of ribosyl-RIF in the experiment described in the supplementary figure. The values for M. flavescens are relatively similar. In addition, the ADP-Ribosyl-RIF is not the final metabolite of the pathway.
In regards of having the entire picture, it is true that we were unable to completely unravel and correlate MIC value, expression of Arr-1, expression of Arr-3, efficiency of each enzyme, production of ADP-Ribosyl-RIF and the presence of other possible mechanisms of resistance and this is indeed a setback in our study, and of most studies ever published, which usually focus on one resistant determinant.
ont guidé mon approche
La première personne du singulier surprend ici, alors que l'article est co-signé.
(re-)médiatisée
Ne devrait-on pas plutôt lire « re-médié », pour faire le lien avec le thème du dossier ?
eLife Assessment
This valuable study used genetic and pharmacological manipulations of insulin/IGF signaling in renal glomerular podocytes to address the role of insulin/IGF axis in podocytes. Solid data are presented to demonstrate that co-inhibition of insulin/IGF signaling in podocytes led to aberrant splicing of mRNAs, which could contribute to the loss of podocytes in vitro and in vivo in mice. As it stands, the study lacks the assessment of developmental phenotype of podocytes in the mouse model.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
Summary:
In this manuscript, the role of the insulin receptor and the insulin growth factor receptor was investigated in podocytes. Mice, where both receptors were deleted, developed glomerular dysfunction and developed proteinuria and glomerulrosclerosis over several months. Because of concerns about incomplete KO, the authors generated and studied podocyte cell lines where both receptors were deleted. Loss of both receptors was highly deleterious with greater than 50% cell death. To elucidate the mechanism of cell death, the authors performed global proteomics and found that spliceosome proteins were downregulated. They confirmed this directly by using long-read sequencing. These results suggest a novel role for insulin and IGF1R signaling in RNA splicing in podocytes.
This is primarily a descriptive study and no technical concerns are raised. The mechanism of how insulin and IGF1 signaling regulates splicing is not directly addressed but implicates potentially the phosphorylation downstream of these receptors. In the revised manuscript, it is shown that the mouse KO is incomplete potentially explaining the slow onset of renal insufficiency. Direct measurement of GFR and serial serum creatinines might also enhance our understanding of progression of disease, proteinuria is a strong sign of renal injury. An attempt to rescue the phenotype by overexpression of SF3B4 would also be useful but may be masked by defects in other spliceosome genes. As insulin and IGF are regulators of metabolism, some assessment of metabolic parameters would be an optional add-on.
Significance:
With the GLP1 agonists providing renal protection, there is great interest in understanding the role of insulin and other incretins in kidney cell biology. It is already known that Insulin and IGFR signaling play important roles in other cells of the kidney. So, there is great interest in understanding these pathways in podocytes. The major advance is that these two pathways appear to have a role in RNA metabolism.
Comments on revised version:
I'm satisfied with the revised manuscript and the responses to my previous concerns.
Reviewer #2 (Public review):
Summary:
In this manuscript, submitted to Review Commons (journal agnostic), Coward and colleagues report on the role of insulin/IGF axis in podocyte gene transcription. They knocked out both the insulin and IGFR1 mice. Dual KO mice manifested a severe phenotype, with albuminuria, glomerulosclerosis, renal failure and death at 4-24 weeks.
Long read RNA sequencing was used to assess splicing events. Podocyte transcripts manifesting intron retention were identified. Dual knock-out podocytes manifested more transcripts with intron retention (18%) compared wild-type controls (18%), with an overlap between experiments of ~30%.
Transcript productivity was also assessed using FLAIR-mark-intron-retention software. Intron retention w seen in 18% of ciDKO podocyte transcripts compared to 14% of wild-type podocyte transcripts (P=0.004), with an overlap between experiments of ~30% (indicating the variability of results with this method). Interestingly, ciDKO podocytes showed downregulation of proteins involved in spliceosome function and RNA processing, as suggested by LC/MS and confirmed by Western blot.
Pladienolide (a spliceosome inhibitor) was cytotoxic to HeLa cells and to mouse podocytes but no toxicity was seen in murine glomerular endothelial cells.
The manuscript is generally clear and well-written. Mouse work was approved in advance. The four figures are generally well-designed, bars/superimposed dot-plots.
Methods are generally well described.
Comments on revised version:
Coward and colleagues have done an excellent job of responding to all the reviewer comments.
Reviewer #4 (Public review):
Summary and background:
This report entitled "The insulin/IGF axis is critically important (for) controlling gene transcription in the podocyte" from Hurcombe et al is based on a mouse double knockdown of the IR and IGF1R and a parallel cultured mouse podocyte model. Insulin/IGF signaling system in mammals evolved as three gene reduplicated peptides (insulin, IGF-1, and IGF-2) and their two receptors IR and IGF1R that cross-react to variable extents with the peptides, are ubiquitously expressed, and signal through parallel pathways. The major downstream effect of insulin is to regulate glucose uptake and metabolism, while that of the IGF pathways is to regulate growth and cell cycling in part through mTORC1. The GH-IGF-1-IGF1R pathway regulates post-natal growth. IGF-2 signaling is thought to play a major role in regulating intrauterine growth and development, although IGF-2 is also present at high levels in post-natal life. Thus, one would anticipate that reducing IR/IGF1R signaling in any cell would slow growth and cell cycling by reducing growth factor and metabolic mTORC1-mediated and other processes including the splicing of RNA for protein synthesis.
Mouse IR/IGF1R double knockdown model:
A double knockdown mouse model was generated by interbreeding mice with different genetic backgrounds carrying floxed sites for IR and IGF-1R to produce mixed background offspring with both floxed IR and IGF-1R genes. These mice were crossed so that the podocin promoter driven-Cre (that comes on at about embryonic day 12 bas podocytes are developing) would delete IR and IGF-1R genes. Since podocin is believed to be an absolutely podocyte-specific protein, this podocin promoter this is predicted to specifically knock down the IR and IGF1R genes only in podocytes. The weight and growth of double KO offspring was not different from controls, but some proportion of the double knockdown mice subsequently developed proteinuria by 6 months and 20% died, although no specific data is provided to identify the cause of the deaths since eGFR was not decreased. Surviving mice were evaluated at 6 months of age. The efficacy of knockdown was not demonstrated in the mouse model itself, although a temperature-sensitive cell line developed from these double knockdown mice showed that expression of IR and IGF-1R proteins in the Cre-treated cell line were both reduced by about 50% (no statistical analysis of this result provided). In the knockout mice, proteinuria was significantly increased by 6 months, but not at earlier time points. Histologic analysis showed proteinaceous casts, glomerulosclerosis and interstitial fibrosis. Podocyte number was stated to be reduced by about 30% in double knockdown mice, although the method by which this was evaluated seems to have been by counting WT1 positive nuclei in glomerular cross-sections, an approach that is well-known not to be a reliable way of assessing true podocyte number. No information is provided about podocyte size, density or glomerular volume.
Comment: If IR/IGF1R deletion plays a significant role in normal podocyte function sufficient to cause proteinuria and glomerulosclerosis then the effect of reduced IR and IGF1R protein expression on podocyte function would have been expected to produce a phenotype before 6 months. A more likely scenario to explain the overall result is that deleting the IR and IGF1R genes at about embryonic day12 impacted podocyte development to a variable extent such that some mice developed fewer podocytes per glomerulus than other mice. As mice grow and their glomeruli and glomerular capillary area increases, those mice with fewer podocytes would not be able to completely cover the filtration surface with foot processes and would develop proteinuria and glomerulosclerosis. If reduced podocyte number per glomerulus is the proximate cause of the observed proteinuria, then modulation of the body and kidney growth rate by calorie restriction to slow growth (lower circulating IGF-1 levels) would be expected to be protective, while a high protein high calorie diet (higher circulating IGF-1 levels) or uni-nephrectomy to increase kidney growth rate would be expected to enhance proteinuria and glomerulosclerosis.
The model as used may be more representative of a variable degree of podocyte depletion than an effect of impaired IR/IGF1R signaling. Therefore, although the phenotype may be ultimately attributable to the IR/IGF1R gene deletions the proteinuria and glomerulosclerotic phenotype itself was probably a consequence of defective podocyte development. Examining podocyte number, size, density and glomerular volume at earlier time points (4 weeks) would help to answer this question. Therefore, a more appropriate title would be "The insulin/IGF axis is critically important (for) normal podocyte development and deployment". In this context the effect of the knockdowns on splicing would make more sense.
Cell culture studies. A cell line was generated using a temperature sensitive SV40 system that has been previously reported from this laboratory. A detailed analysis is provided to show that double knockout cells exhibited abnormal spliceosome activity. This forms the basis for the conclusion that "The insulin/IGF axis is critically important (for) controlling gene transcription in the podocyte". There are several concerns that weaken this conclusion.
(1) In the double knockdown cell culture system about 30% of cells were "lost" by 3 days and about 70% of cells were "lost" by 5days. The studies were done at the 3 day time point. It is not clear whether "lost" cells were in the process of dying, stress-induced detachment, or just growing more slowly than control due to reduced IR and IGF-1R signaling. These processes could have impacted splicing in a non-specific way independent of IR/IGF1R signaling itself.
(2) Can a single cell line derived from the double floxed mice be relied on to provide an unbiased picture of the effect of deleting IR and IGF-1R? Presumably, the transfection and selection process will select for cells that survive thereby including unknown biases, possibly related to spliceosome function. Is a single cell line adequate? These investigators have extensive experience with this type of analysis, but this question is not addressed in the discussion.
(3) To determine whether the effect is specific to reduced IR/IGFR signaling the deletion of IR and IGF-1R could be corrected by transfecting full length IR and IGF-1R cDNAs into the cells to restore normal IR/IGF1R signaling. If transfected cells with intact IR and IGF-1R expression and activity returns spliceosome activity to normal this would be evidence that receptors themselves play some role in spliceosome activity, as opposed to the downstream effect on growth limitation/stress on the cells.
(4) Other ways of testing whether the splicing effect is specifically due to reduced IR/IGF-1R signaling would be to (a) block IR and IGF1R receptors using available inhibitors, (b) remove or reduce insulin, IGF-1 and IGF-2 levels in the culture medium, (c) use low glucose and amino acid culture medium to slow growth rate independent of receptor function, (d) or block intra-cellular signaling via the IR and IGF-1R receptors through mTORC1 inhibition using rapamycin or other signaling targets.
(5) It would be useful to determine whether the cultured cells stressed in other ways (e.g. ischemia, toxins, etc.) also results in the same splicing abnormalities.
Author response:
The following is the authors’ response to the original reviews
Many thanks for your helpful and constructive comments for our work examining the effect of inhibiting both the insulin receptor (IR) and IGF1 receptor (IGF1R) in the podocyte. We are pleased to submit an updated manuscript addressing your concerns.
(1) A major concern was a lack of mechanistic insight into how deletion (or knock-down) of both receptors caused the spliceosomal phenotype (Reviewer 1 and Reviewer 3).
We now think this is due to the lack of a network of insulin/IGF phospho-signalling events to a variety of spliceosomal proteins and kinases. The reasons for this are as follows:
A. Since submitting our paper Turewicz et al have published a comprehensive phospho-proteomic paper examining the effects of 100nM insulin on human primary myotubes (DOI: 10.1038/s41467-025-56335-6). They discovered that multiple post-translational phosphorylation events occur in a variety of spliceosomal proteins at differing time points (1 minute to 60 minutes). Furthermore, they show that mRNA splicing is rapidly modified in response to insulin stimulation in their cells. This follows elegant work from Bastista et al who studied diabetic and non-diabetic iPSC derived human myositis and also detected a spliceosome phosphorylation signature (DOI: 10.1016/j.cmet.2020.08.007).
B. We have examined phospho-proteosome changes that occur in wild -type podocytes (expressing both the IR and IGF1R) compared to double (IR and IGF1R) knockout cells using phosho-proteomics. We have done this 3 days after inducing receptor knockdown, before major cell loss, and have stimulated the cells with either 10nM insulin or 100mg IGF1.
Interestingly, we detected several post-translational modifications (PTM) in our data set that are also present in Turewicz’s studies. Of note, 100nM insulin (as used by Turewicz) will signal through both the insulin and IGF1 receptor (and hybrid Insulin/IGF1 receptors) which is relevant to our studies.
Our work shows a cascade of phospho- signalling events affecting multiple components of the spliceosomal complex and evidence of kinase modulation (phosphorylation) (New Figure 7 and supplementary Figure 5). Also new results section in paper (lines 391-425 in track changes version). We acknowledge that we only studied a single time point after stimulation (10 minutes) and could have missed other PTM in the spliceosomal complex and other kinases. This is mentioned in our new limitations of study section (lines 595-606). This will be a focus of future work. We did not find major PTM differences when stimulating with either insulin or IGF1 in our studies and suspect that the doses of insulin (10nM) and IGF1 (100mg) used are still able to signal through cognate receptors.
Furthermore, we have examined the relative contributions of the insulin and IGF1 receptor in detail in the model (addressed in point 13 below).
(2) The phenotype of the mouse is only superficially addressed. The main issues are that the completeness of the mouse KO is never assessed nor is the completeness of the KO in cell lines. The absence of this data is a significant weakness. (Reviewer 1)
We apologise for not making this clear, but we did assess the level of receptor knockdown in both the animal and cell models. The in vivo model showed variable and non-complete levels of insulin receptor and IGF1 receptor podocyte knock down (shown in supplementary Figure 1C). This is why we made the in vitro floxed podocyte cell lines in which we could robustly knockdown both the IR and IGF1R. We show this using Western blotting (shown in Figure 2A). We agree that calling the models knockout is misleading and have changed all to knock down (KD) now.
(3) The mouse experiments would be improved if the serum creatinine’s were measured to provide some idea how severe the kidney injury is. (Reviewer 1)
There is variability in creatinine levels which is not uncommon in transgenic mouse models (probably partly due to variability in receptor knock down levels with cre-lox system). This is part of rationale of developing the robust double receptor knockout cell models where we robustly knocked out both receptors by >80%. We have added measured creatinine levels in a subset of mice in supplementary data (New Supplementary Figure 1E) and mention this in the text (lines 285-286). As some mice died we expect they may have developed acute kidney injury, but we did not serially measure the creatinine’s in every mouse over time. We could have assessed the GFR in a more sensitive way to look at differences. However, we consider the highly significant levels of albuminuria and histological damage observed in our models show a significant kidney phenotype.
(4) An attempt to rescue the phenotype by overexpression of SF3B4 would also be useful. If this didn't work, an explanation in the text would suffice. (Reviewer 1).
We did consider doing this but on reflection think it is very unlikely to rescue the phenotype as an array of different spliceosomal proteins quantitatively changed and were differentially phosphorylated / dephosphorylated throughout the complex (as we hope our revised work illustrates now). We think a single protein rescue is highly unlikely to work. We hope this is an appropriate explanation for this action. We have mentioned this in the text now in our discussion (lines 601-602).
(5) As insulin and IGF are regulators of metabolism, some assessment of metabolic parameters would be an optional add-on. (Reviewer 1).
Thank you for this suggestion. We did not extensively examine the metabolism of the mice however we did perform blood glucose measurement and weight which are included in the paper (Figure 1A and Figure 1B).
(6) The authors should caveat the cell experiments by discussing the ramifications of studying the 50% of the cells that survive vs the ones that died. (Reviewer 1).
We appreciate this and this was the rationale behind cells being studied after 3 days differentiation for total and phospho-proteomics before significant cell loss to avoid the issue of studying the 50% of cells that survive (which happened at 7 days). We have made this clearer in the manuscript. We also have added the data showing less cell death at 3 days in the cell model (New Supp Figure 2B).
(7) It would be helpful to say that tissue scoring was performed by an investigator masked to sample identity. (Reviewer 2)
We did this and have added to manuscript (line 113).
(8) Data are presented as mean/SEM. In general, mean/SD or median/IQR are preferred to allow the reader to evaluate the spread of the data. There may be exceptions where only SEM is reasonable. (Reviewer 2)
All graphs have now been changed to SD rather than SEM.
(9) It would be useful to for the reader to be told the number of over-lapping genes (with similar expression between mouse groups) and the results of a statistical test comparing WT and KO mice. The overlap of intron retention events between experimental repeats was about 30% in both knock-out podocytes. This seems low and I am curious to know whether this is typical for this method; a reference could be helpful. (Reviewer 2)
This is an excellent question. We had 30% overlap as the parameters used for analysis were very stringent. We suspect we could get more than 30% by being less stringent, which still be considered as similar events if requested. Our methods were based on FLAIR analysis (PMID: 32188845). We have added this reference to the manuscript (Line 242 & 680).
(10) With the GLP1 agonists providing renal protection, there is great interest in understanding the role of insulin and other incretins in kidney cell biology. It is already known that Insulin and IGFR signaling play important roles in other cells of the kidney. So, there is great interest in understanding these pathways in podocytes. The major advance is that these two pathways appear to have a role in RNA metabolism, the major limitations are the lack of information regarding the completeness of the KO's. If, for example, they can determine that in the mice, the KO is complete, that the GFR is relatively normal, then the phenotype they describe is relatively mild. (Reviewer 1)
Thank you. The receptor knock-out (KO) in the mice is highly unlikely to be complete (Please see comments above and Supplementary Figure 1C). There are many examples of “KO” animal models targeting other tissues showing that complete KO of these receptors seems difficult to achieve, particularly in reference to the IGF1 receptor. In the brain, which also contains terminally differentiated cells, barely 50% of IGF1R knockdown was achieved in the target cells (PMID:28595357). In ovarian granulosa cells (PMID:28407051) -several tissue specific drivers tried but couldn't achieve any better than 80%. The paper states that 10% of IGF1R is sufficient for function in these cells so they conclude that their knockdown animals are probably still responding to IGF1. Finally, in our recent IGF1R podocyte knockdown model we found Cre levels were important for excision of a single homozygous floxed gene (PMID: 38706850) hence we were not surprised that trying to excise two homozygous floxed genes (insulin receptor and IGF1 receptor) was challenging. This was the rationale for making the double receptor knockout cell lines to understand processes / biology in more detail. As stated earlier, we have changed our description of the mice and cell lines from knock-out to knock-down throughout the revised manuscript as this is more accurate.
(11) For the in vivo studies, the only information given is for mice at 24 weeks of age. There needs to be a full-time course of when the albuminuria was first seen and the rate of development. Also, GFR was not measured. Since the podocin-Cre utilized was not inducible, there should be a determination of whether there was a developmental defect in glomeruli or podocytes. Were there any differences in wither prenatal post-natal development or number of glomeruli? (Reviewer 3)
We have added further urinary Albumin:creatinine ratio (uACR) data at 12, 16 and 20 weeks to manuscript. We do not think there was a major developmental phenotype as albuminuria did not become significantly different until several months of age (new Supp Figure 1B). We did consider using a doxycycline inducible model but we know the excision efficiency is much less than the constitutive podocin-cre driven model Author response image 1. This would likely give a very mild (if any) phenotype when attempting to knockout both receptors and not reveal the biology adequately. We acknowledge the weaknesses of the animal model and this was the rationale for generating the cell models.
(12) Although the in vitro studies are of interest, there are no studies to determine if this is the underlying mechanism for the in vivo abnormalities seen in the mice. Cultured podocytes may not necessarily reflect what is occurring in podocytes in vivo. (Reviewer 3)
This is a good point. We have now immune-stained the DKD and WT mice for Sf3b4 (a spliceosomal change in our in vitro proteomics) and also find a significant reduction in this protein in podocytes of the DKD mice (New Figure 3F).
(13) Given that both receptors are deleted in the podocyte cell line, it is not clear if the spliceosome defect requires deletion of both receptors or if there is redundancy in the effect. The studies need to be repeated in podocyte cell lines with either IR or IGFR single deletions. (Reviewer 3)
We have now performed proteomics and phospho-proteomics in all 4 cell types (Wild-type, Insulin receptor knock down, IGF1R knockdown and double knockdown) at 3 days (New Figure 8 and supplementary Figure 6. Also new results section lines 425 to 450). This shows that both receptors contribute to the pathways (and hence there is a high level of compensation built into the system). For total proteins we detected that spliceosomal tri-snRNP was only reduced when both receptors were lacking but other proteins / pathways had an incremental effect of losing the insulin or IGF1 receptor. Likewise, the spliceosomal phospho-signaling events can go through either the insulin or igf1 receptors predominantly or through both. We think this reflects the complexity of this system and how evolutioatily it has developed in mammals to protect against its loss.
Finally in revision we have rewritten the discussion with a “limitations of the study” section and hopefully in an easier to read fashion for the readership.
Author response image 1.
(A) mT/mG reporter mouse crossed to constitutional podocin Cre heterozygous mouse. Illustrates podocyte specificity for Cre driver and excision Of reporter Figure shows GFP expression in Cre producing cells (top panel scale bar=250vm; bottom panel scale bar=50pm). Cre expression causes GFP to be switched on. (B) mT/mG reporter mouse crossed to podocin RtTA— tet-o-cre heterozygous mouse shows podocyte specificity for driver and approximately 60% excision. (top and bottom panels scale bar=250pm; middle panel scale bar=50pm). Doxycycline required for expression showing not leaky.
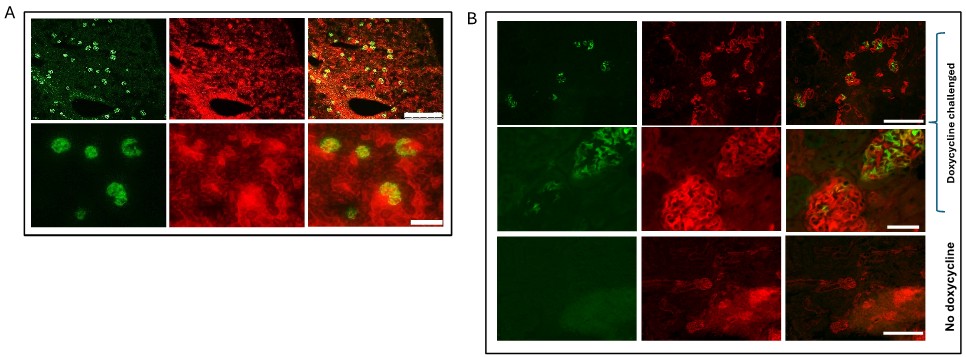
L'Exclusion Scolaire et la Fabrique du Décrochage : Enjeux et Mécanismes
L'exclusion d'un élève, qu'elle soit ponctuelle ou définitive, est devenue un fait banalisé dans le système éducatif français.
Pourtant, les recherches de Julien Garric, enseignant-chercheur en sciences de l'éducation, révèlent que cette pratique constitue un levier majeur du décrochage scolaire.
La France se distingue par une culture de la sanction particulièrement marquée, facilitée par une structure unique : le service de "Vie scolaire".
Ce système permet une externalisation rapide des conflits de classe, mais crée un cercle vicieux où l'élève, de plus en plus éloigné des apprentissages, finit par intérioriser son exclusion.
Le manque de données nationales et de formation sur le sujet occulte une réalité sociale brutale : le décrochage touche de manière disproportionnée les garçons et les élèves issus de milieux défavorisés (REP+), transformant l'école en un lieu où la réussite des uns semble parfois dépendre de l'éviction des plus fragiles.
--------------------------------------------------------------------------------
Le système éducatif français présente des caractéristiques uniques en Europe concernant la gestion des comportements des élèves.
• L'appel d'air de la "Vie Scolaire" : Contrairement à ses voisins européens, l'établissement français dispose d'un service dédié (CPE, AED) qui permet de prendre en charge l'élève en dehors de la salle de classe. Cette existence d'un "ailleurs" facilite l'éviction ponctuelle des élèves perturbateurs.
• Externalisation du traitement : Cette structure permet aux enseignants du secondaire de ne pas avoir à gérer seuls les difficultés de comportement, à l'inverse des enseignants du primaire ou de leurs homologues européens qui doivent trouver des solutions internes à la classe.
• Une culture punitive forte : Les enquêtes de comparaison internationale suggèrent que la France punit davantage et plus sévèrement que les autres pays européens.
Ce recours massif à la sanction (exclusion de cours, retenues, exclusions temporaires ou définitives) crée un paradoxe : les enseignants réclament plus de sévérité alors que les élèves français ressentent un sentiment d'injustice plus fort que partout ailleurs en Europe.
Malgré l'existence d'outils numériques comme Pronote qui recensent chaque punition, le phénomène de l'exclusion reste largement sous-étudié et peu quantifié à l'échelle nationale.
| Type de donnée | État des lieux | | --- | --- | | Données locales (Pronote) | Mine d'informations inexploités au niveau global ; recense les exclusions formelles. | | Pratiques informelles | Non quantifiées (élèves laissés dans le couloir ou chez un voisin). | | Communications institutionnelles | Absence de notes d'information régulières de la DEPP sur les sanctions et les conseils de discipline. | | Formation des personnels | Sujet largement absent de la formation initiale et continue des enseignants. |
Cette absence de volonté politique de centraliser les données suggère un désintérêt pour une pratique jugée "peu glorieuse" et souvent réduite, dans les rapports d'inspection, à une responsabilité individuelle du personnel.
Les recherches mettent en évidence une corrélation forte entre les politiques de "tolérance zéro" et l'augmentation de la violence et du décrochage.
1. Le cercle vicieux de l'apprentissage : L'élève exclu passe plus de temps en Vie scolaire qu'en classe. Cet éloignement physique des cours aggrave ses difficultés scolaires initiales, le rendant encore moins apte à suivre les enseignements à son retour.
2. La rupture relationnelle : L'exclusion répétée provoque une escalade dans le conflit. L'élève se sent rejeté et construit des comportements de plus en plus déviants, tandis que l'enseignant, à bout de ressources, multiplie les évictions.
3. Le paradoxe de la Vie scolaire : Pour certains élèves fragiles, la Vie scolaire devient un refuge où ils trouvent l'écoute et le réconfort qu'ils ne trouvent plus en classe.
Cependant, ce réconfort les éloigne définitivement du cœur de la mission scolaire : l'acquisition de connaissances.
L'exclusion ne frappe pas au hasard ; elle suit des lignes de fracture sociales et de genre très nettes.
• Le genre (80 % de garçons) : Les garçons sont massivement plus punis.
Ce phénomène repose sur des stéréotypes de genre ancrés chez les adultes (parents et personnels), qui attendent des garçons des comportements plus agités.
En réaction, certains garçons ne trouvant pas de valorisation par les notes cherchent une forme de reconnaissance perverse à travers la punition.
• Le déterminisme social :
◦ Familles les plus favorisées : moins de 1 % de décrochage.
◦ Éducation prioritaire renforcée (REP+) : plus d'un tiers des élèves décrochent.
• Le destin scolaire anticipé : Dans les quartiers les plus pauvres, le décrochage est un aboutissement courant.
Dès la classe de sixième, certains élèves font le "deuil" d'une scolarité longue et intériorisent qu'ils ne sont pas des élèves comme les autres.
L'acte d'exclure n'est jamais anodin et produit des effets délétères sur l'ensemble de la communauté éducative.
• Pour l'élève exclu : Une attaque violente contre l'estime de soi.
L'exclusion est une désignation publique devant les pairs qui force l'élève à adopter des stratégies pour "sauver la face", souvent par le défi ou l'indifférence feinte.
• Pour les enseignants : Un sentiment de souffrance et de contradiction avec leurs propres valeurs.
Personne ne s'engage dans l'enseignement pour "mettre des élèves à la porte", mais l'absence de pratiques collectives alternatives mène à ces situations par défaut.
• Pour les élèves non-exclus : Une éducation à la citoyenneté paradoxale où les "vainqueurs" du système assistent, impuissants ou spectateurs, à l'élimination progressive des "perdants" (les plus fragiles).
Pour sortir de l'impasse, le passage d'une sanction individuelle à une réflexion d'équipe est nécessaire.
• Le "moment du retour" : La faille majeure de l'exclusion réside dans l'absence de médiation lors du retour de l'élève.
La punition ne devient éducative que si elle est suivie d'un temps de réparation où l'enseignant et l'élève reconstruisent le lien cassé.
• Le théorème de la protection du groupe : Il faut déconstruire l'idée que le sacrifice d'une minorité (1 ou 2 élèves par classe) est le seul moyen de protéger la majorité.
Cumulés à l'échelle d'un collège, ces "sacrifices" constituent une part massive de la jeunesse exclue du système de certification.
• Apprentissage des codes : Les normes de comportement font partie du "curriculum caché". L'école doit enseigner explicitement comment se comporter, au lieu de sanctionner des élèves qui tâtonnent et ne possèdent pas les codes implicites attendus au collège.
Citation clé : « On peut travailler autant qu'on veut sur l'amélioration des savoirs fondamentaux, si un certain nombre d'élèves n'assistent pas aux cours parce que le système pense qu'ils n'y ont pas leur place, on n'y arrivera pas. » — Julien Garric
To crave a passport for my wand’ring Ghost:
after his death he was lost, he did not go to heaven nor hell. Him being lost and trapped means he has not found peace = revenge being suggested.
Did live
immediately indicate that the character speaking on stage is clearly dead. So, the audience now knows that they have been introduced to a ghost.
The Barbell Method is a phrase coined by Nassim Taleb. It means that you make sure that the majority of your investment is safe while you make small but very risky bets. You combine safety with the possibility of high revenue. The trick is that you floor the possible loss while leaving the possible revenue unlimited.
gali tikti bet kam, kai reikia rinktis iš daug ir/ar skirtingos rizikos pasirikimų
Analyse du Binôme de Direction en Milieu Scolaire : Vers un Modèle de Coresponsabilité
Ce document analyse les dynamiques complexes au sein du binôme de direction (chef d’établissement et adjoint) dans le système éducatif français.
Fondé sur les recherches de Rosenne Descré Rouillard, il met en lumière l'obsolescence du modèle traditionnel pyramidal qui conduit souvent à l'épuisement du dirigeant et à la frustration de l'adjoint.
L'analyse révèle que le binôme fonctionne comme un « couple forcé » où l'intime et le professionnel s'entremêlent, rendant la relation soit extrêmement puissante, soit pathogène.
Pour transformer cette tension en partenariat efficace, il est impératif de passer d'une répartition des tâches subie à une coresponsabilité basée sur la confiance, la transparence et la reconnaissance des compétences individuelles.
--------------------------------------------------------------------------------
Le modèle classique de direction est marqué par une division du travail héritée et sociale, bien que non inscrite officiellement dans les textes.
• La répartition hiérarchique : Traditionnellement, le chef d'établissement conserve le pilotage stratégique et pédagogique, tandis que l'adjoint est cantonné à l'organisationnel, au technique et à l'exécution.
• Le « sale boulot » : Les recherches décrivent l'adjoint comme un « artisan du quotidien » occupant une fonction intervalle.
Il récupère souvent les tâches les moins valorisées et les plus invisibles, ce que la sociologie qualifie de « sale boulot ».
• L’asymétrie de fonction : Bien qu'appartenant au même corps de métier (personnel de direction), l'adjoint doit rester en « seconde cordée » ou agir comme un « copilote », ce qui crée un décalage entre sa formation de chef et sa réalité opérationnelle.
• Charge mentale explosive : Le chef d'établissement, seul responsable légal et comptable, subit une pression qui freine la délégation.
• Sous-utilisation des compétences : L'adjoint peut ressentir une frustration légitime lorsque ses compétences stratégiques sont ignorées au profit d'une gestion purement logistique.
• Atterrissage brutal : Pour beaucoup de nouveaux adjoints, le passage du concours à la réalité du terrain est vécu comme un choc, car ils sont formés pour diriger mais se retrouvent en position subalterne.
--------------------------------------------------------------------------------
La relation entre le chef et l'adjoint dépasse le simple cadre fonctionnel pour toucher à la sphère de l'intime.
• L'absence de choix : Les membres du binôme ne se choisissent pas.
Cette union imposée par l'institution crée une « épreuve bicéphale » où partager le pouvoir et l'autorité devient un défi quotidien.
• L'isolement à deux : Contrairement aux enseignants ou aux CPE qui travaillent en communauté, le binôme de direction est souvent isolé.
Cette solitude partagée renforce la nécessité d'une entente parfaite.
• L'impact de la personnalité : Quand le binôme « matche », il devient une force extrême.
Quand il « clashe », cela peut mener à des maladies professionnelles tant l'implication personnelle est forte.
Le binôme doit « parler d'une seule voix » devant la communauté éducative (enseignants, élèves, parents), à l'instar d'un couple de parents devant ses enfants.
Les désaccords doivent être réglés en privé pour éviter que les tiers ne s'engouffrent dans les failles de la direction.
--------------------------------------------------------------------------------
Pour sortir des tensions, le binôme doit instaurer un modèle de coresponsabilité.
| Pilier | Description et Mise en Œuvre | | --- | --- | | Loyauté et Confiance | Socle indispensable qui doit être total et réciproque pour permettre au binôme de « faire front » face aux pressions institutionnelles. | | Transparence Absolue | Partage intégral des informations pour qu'aucun membre ne soit pris au dépourvu. | | Complémentarité | S'appuyer sur les appétences et les métiers d'origine (ex: un ancien CPE sur le leadership éducatif, un enseignant sur la pédagogie). | | Unité de Façade | Adopter une position commune indéfectible à l'extérieur, même si les tonalités de voix diffèrent. | | Égalité de Coopération | Considérer l'adjoint comme un véritable partenaire d'égal à égal plutôt que comme un « super secrétaire ». |
--------------------------------------------------------------------------------
La transition vers une direction partagée nécessite des actions concrètes et des rituels de communication.
• Délégation complète : Le chef doit autoriser l'adjoint à gérer des dossiers de A à Z sans intervenir de manière intempestive, favorisant ainsi l'autonomie et la montée en compétences.
• Lettre de mission évolutive : Cet outil doit être coconstruit et révisé à chaque changement de binôme pour refléter les compétences réelles et non une répartition automatique.
• Interchangeabilité : Dans un binôme fluide, chaque membre doit être capable de prendre le relais sur les dossiers de l'autre en cas d'absence.
• Échanges informels quotidiens : Maintenir une politique de « bureau porte ouverte » pour une interconnexion permanente.
• Le point hebdomadaire : Se réserver un temps dédié (par exemple le vendredi soir) pour « rembobiner le fil de la semaine », analyser les pratiques et évacuer les tensions.
• Protection mutuelle : Le chef a un rôle de protecteur ultime envers l'adjoint, mais l'adjoint doit aussi veiller sur le chef.
• Balises horaires : S'imposer des limites mutuelles sur le temps de travail et l'usage du numérique pour prévenir l'épuisement, particulièrement complexe dans le cadre des logements de fonction.
--------------------------------------------------------------------------------
L'analyse conclut que l'institution doit évoluer pour soutenir ces nouvelles formes de gouvernance.
• Vers la coresponsabilité légale : Une évolution législative pourrait instaurer une véritable codirection, soulageant ainsi la responsabilité unique du chef.
• Amélioration des RH : Dépasser les règles d'ancienneté pour former des binômes basés sur la complémentarité des profils.
• Formation continue : Intégrer la gestion de la relation humaine et du binôme dès la préparation au concours pour éviter « l'atterrissage brutal ».
• Espaces de parole : Développer des temps d'analyse de pratique ou de coaching collectif, extérieurs à la hiérarchie, pour permettre aux personnels de direction de se « resocialiser professionnellement » à chaque changement de partenaire.
En résumé, le succès d'un binôme de direction repose sur sa capacité à transformer une hiérarchie rigide en un management horizontal partagé, où l'humain est placé au centre de la stratégie de pilotage.
trophies, the effect of these mechan-ics becoming so solidly integrated into videogame culture is that theirgravitational pull changes how games are played and interpreted. Lea-derboards are a clear place where players are compared with one an-other. A former highly competitive Madden NFL player writes thatleaderboards are “a devilish feature,” as they transform “Madden froman escapist pastime into another stage on which to prove your self-worth.”36 Gamerscore is a measure that turns abstract effort in a gameinto concrete results that are intelligible to others at a mere glimpse.
I 110% this hard game and you didn't heh...
Theindividual impact on your rating within LoL fosters the perceptionamong some of an “Elo hell” that is “populated by griefers/trolls and‘bad’ players that prevent them from moving up the ranks.”13 A guideto ranked games in LoL contends that Elo hell is a figment of players’imaginations, since “a large misconception is that it is always team-mates that is bringing you down [sic],” and the answer is that “ratingsbecome more accurate the more games are played [sic]. Think ‘big pic-ture.’ It can take hundreds or thousands of games to be consistentlymatched with similarly skilled players.”14 Although incredibly rationaland likely true, this kind of thinking is much harder to remember inthe moment when you believe some other player has cost you some-thing through his or her inactivity or poor performance.
And who is fighting to go to the World Championships? Much like in mainstream sports, white (or asian), young, rich, males.
The process turned participation in desired activities intopoints that could then be spent on rewards. Silverman and Simon dis-cuss how some of the best guilds in the world rejected DKP becauseit focuses players on rewards, rather than on group accomplishments
Campbell's or Goodhart's laws: The more any quantitative social indicator is used for social decision-making, the more subject it will be to corruption pressures and the more apt it will be to distort and corrupt the social processes it is intended to monitor.
In other words, positivist metricism; which is a form of snob credentialism.
A hexagon map editor for fantasy mapping as plugin for Obsidian. Vgl fantasy cartography at CaL 2019, https://www.zylstra.org/blog/2019/06/getting-frodo-to-mordor-with-geo-data-analysis/
Gestion des élèves perturbateurs : approches psychopédagogiques et cadres éthiques
Le comportement perturbateur d'un élève ne doit pas être perçu comme une simple transgression, mais comme le symptôme d'un mal-être profond, souvent enraciné dans un vécu personnel ou scolaire difficile.
La gestion efficace de ces situations repose sur la reconnaissance des besoins psychologiques fondamentaux de l'élève (sécurité, reconnaissance, justice, estime de soi) et sur la mise en place d'espaces de parole institutionnalisés.
L'analyse souligne que de nombreux élèves perturbateurs, y compris les harceleurs, sont eux-mêmes en situation de souffrance.
Pour répondre à ces défis, les personnels de direction et les équipes éducatives doivent naviguer entre quatre orientations éthiques :
L'équilibre entre ces dimensions permet de maintenir le lien de confiance entre l'élève et l'institution, évitant ainsi le décrochage ou l'exclusion définitive des profils les plus vulnérables.
Le comportement perturbateur est défini comme une manifestation de symptômes liés à une insatisfaction des besoins psychologiques fondamentaux.
Aider un élève nécessite d'être attentif à ces signes, qu'ils soient émotionnels ou plus subtils.
Pour remédier aux comportements problématiques, l'institution doit prendre en considération :
L'intervention repose sur une distinction claire entre les problématiques collectives et individuelles, ainsi que sur la création de structures d'échange formelles.
• Problème collectif : Doit faire l'objet d'une discussion collective.
• Problème individuel : Le comportement d'un élève spécifique doit être traité exclusivement avec lui, afin de préserver sa dignité et de favoriser un dialogue constructif.
La mise en place de "cellules d'écoute" ou d'espaces de parole sécurisés est présentée comme une solution aux résultats rapides et significatifs.
• L'écoute active : Les adultes doivent être formés pour permettre à l'élève d'élaborer lui-même le sens de son vécu.
• Efficacité constatée : Des exemples, notamment dans l'académie de Grenoble, montrent qu'une participation à deux ou trois reprises à ces espaces peut transformer le comportement des jeunes.
• Sécurité affective : L'espace doit permettre à l'élève de dire ce qu'il ressent sans crainte immédiate de jugement ou de répression.
Face à des actes graves (comme des injures envers un enseignant), la sanction demeure nécessaire.
Cependant, la procédure doit respecter des principes éthiques et réglementaires stricts.
• Le principe du contradictoire : Avant et pendant le conseil de discipline, toutes les parties doivent pouvoir s'exprimer et clarifier les faits.
• L'analyse de la souffrance : Il est impératif de considérer que l'élève auteur d'actes délictueux est souvent un élève qui souffre.
Le document note par exemple qu'un grand nombre de harceleurs sont eux-mêmes victimes de harcèlement.
• Dialogue avec la famille : La compréhension du contexte familial est cruciale pour identifier les racines du comportement de l'adolescent.
Le chef d'établissement et son équipe doivent composer avec quatre dimensions éthiques lors de la prise de décision disciplinaire :
| Orientation éthique | Définition et application | | --- | --- | | Déontologique | Respect strict du règlement intérieur et de la loi. C'est l'approche systématique : "à tel acte correspond telle sanction". Essentiel pour la responsabilité professionnelle du chef d'établissement. | | Conséquentialiste | Attention portée aux conséquences de la sanction sur l'avenir de l'élève. Par exemple, éviter d'informer des parents violents d'une faute mineure pour ne pas infliger une "double peine" à l'enfant. | | Exercice des vertus | Mise sur la patience, la prudence et la confiance. On donne du temps à l'élève pour s'améliorer en privilégiant un blâme ou un avertissement plutôt qu'une exclusion. | | Éthique du Care (Soin) | Posture indispensable vis-à-vis des élèves les plus vulnérables traversant des souffrances psychiques graves. Il s'agit de maintenir la "tête hors de l'eau" pour l'élève par un regard attentif et bienveillant. |
Le chef d'établissement a la responsabilité première de garantir le respect de la règle et du droit (réflexe déontologique) pour éviter toute faute professionnelle. Toutefois, la réalité du terrain impose une composition entre ces différentes éthiques.
Une décision efficace est souvent hybride : elle rappelle la règle (déontologie), tout en tempérant la sanction au regard du contexte (conséquentialisme) et en demandant à l'équipe pédagogique une "bienveillance attentive" (care).
Cette approche intégrée est présentée comme le seul moyen de préserver la confiance des élèves les plus fragiles envers l'école et les adultes, prévenant ainsi leur exclusion définitive du système scolaire.
eLife Assessment
In this valuable study, through carefully executed and rigorously controlled experiments, the authors challenged a previously reported role of the Death Receptor 6 (DR6/Tnfrsf21) in Wallerian degeneration (WD). Using two DR6 knockout mouse lines and multiple WD assays, both in vitro and in vivo, the authors provided convincing evidence that loss of DR6 in mice does not protect peripheral axons from WD after injury, at least in the specific contexts of the mice and analyses performed in this study. Due to the lack of certain specific parameters from previous studies (sex, age, mouse strains etc.), the exact reasons underlying the observed inconsistencies between current and previous reports on the protective effects of DR6 remains to be determined. Overall, this is a carefully executed study providing invaluable information toward understanding DR6's role (or lack thereof) in axon degeneration.
Reviewer #1 (Public review):
Summary:
The authors show that genetic deletion of the orphan tumor necrosis factor receptor DR6 in mice does not protect peripheral axons against degeneration after axotomy. Similarly, Schwann cells in DR6 mutant mice react to axotomy similarly to wild type controls. These negative results are important because previous work has indicated that loss or inhibition of DR6 is protective in disease models and also against Wallerian degeneration of axons following injury. This carefully executed counterexample is important for the field to consider.
Strengths:
A strength of the paper is the use of two independent mouse strains that knockout DR6 in slightly different ways. The authors confirm that DR6 mRNA is absent in these models (western blots for DR6 protein are less convincingly null, but given the absence of mRNA, this is likely an issue of antibody specificity). One of the DR6 knockout strains used is the same strain used in a previous paper examining the effects of DR6 on Wallerian degeneration.
The authors use a series of established assays to evaluate axon degeneration, including light and electron microscopy on nerve histological samples and cultured dorsal root ganglion neurons in which axons are mechanically severed and degeneration is scored in time lapse microscopy. These assays consistently show a lack of effect of loss of DR6 on Wallerian degeneration in both mouse strains examined.
Additional strengths are that the authors examine both the axonal response and the Schwann cell response to axotomy and use both in vivo and in vitro assays.
Therefore, these experiments, the author's data support their conclusion that loss of DR6 does not protect against Wallerian degeneration.
Weaknesses:
A weakness of this paper is that no effort is made to determine why the results presented here may differ from previous studies. A notable possibility is that the original mouse strain that showed 5 of 13 mice being protected from Wallerian degeneration was studies on a segregating C57BL/6.129S background.
Finally, it is important to note that previously reported effects of DR6 inhibition, such as protection of cultured cortical neurons from beta-amyloid toxicity, are not necessarily the same as Wallerian degeneration of axons distal to an injury studied here. The negative results presented here showing that loss of DR6 is not protective against Wallerian degeneration induced by injury are important given the interest in DR6 as a therapeutic target. However, care should be taken in attempting to extrapolate these results to other disease contexts such as ALS or Alzheimer's disease.
Reviewer #3 (Public review):
Summary:
The authors revisit the role of DR6 in axon degeneration following physical injury (Wallerian degeneration), examining both its effects on axons and its role in regulating the Schwann cell response to injury. Surprisingly, and in contrast to previous studies, they find that DR6 deletion does not delay the rate of axon degeneration after injury, suggesting that DR6 is not a mediator of this process.
Overall, this is a valuable study. As the authors note, the current literature on DR6 is inconsistent, and these results provide useful new data and clarification. This work will help other researchers interpret their own data and re-evaluate studies related to DR6 and axon degeneration.
Strengths:
(1) The use of two independent DR6 knockout mouse models strengthens the conclusions, particularly when reporting the absence of a phenotype.
(2) The focus on early time points after injury addresses a key limitation of previous studies. This approach reduces the risk of missing subtle protective phenotypes and avoids confounding results with regenerating axons at later time points after axotomy.
Comments on revisions:
I thank the authors for their thorough responses to my previous comments. The revisions have addressed the points raised and have improved the clarity and overall quality of the manuscript. I appreciate the effort taken to strengthen the presentation of the work.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors show that genetic deletion of the orphan tumor necrosis factor receptor DR6 in mice does not protect peripheral axons against degeneration after axotomy. Similarly, Schwann cells in DR6 mutant mice react to axotomy similarly to wild-type controls. These negative results are important because previous work has indicated that loss or inhibition of DR6 is protective in disease models and also against Wallerian degeneration of axons following injury. This carefully executed counterexample is important for the field to consider.
Strengths:
A strength of the paper is the use of two independent mouse strains that knock out DR6 in slightly different ways. The authors confirm that DR6 mRNA is absent in these models (western blots for DR6 protein are less convincingly null, but given the absence of mRNA, this is likely an issue of antibody specificity). One of the DR6 knockout strains used is the same strain used in a previous paper examining the effects of DR6 on Wallerian degeneration.
The authors use a series of established assays to evaluate axon degeneration, including light and electron microscopy on nerve histological samples and cultured dorsal root ganglion neurons in which axons are mechanically severed and degeneration is scored in time-lapse microscopy. These assays consistently show a lack of effect of loss of DR6 on Wallerian degeneration in both mouse strains examined.
Therefore, in the specific context of these experiments, the author's data support their conclusion that loss of DR6 does not protect against Wallerian degeneration.
Weaknesses:
(1) The major weaknesses of this paper include the tone of correcting previously erroneous results and the lack of reporting on important details around animal experiments that would help determine whether the results here really are discordant with previous studies, and if so, why.
The authors do not report the genetic strain background of the mice used, the sex distributions of their experimental cohorts, or the age of the mice at the time the experiments were performed. All of these are important variables.
(Response 1) We thank the reviewer for emphasizing the importance of reporting the sex, age, and genetic background of the experimental animals used in our axon protection analyses. We have incorporated this information into the revised manuscript wherever available. The sole exception concerns the genetic background of the conditional DR6 mice generated by Genentech, which remains unknown. The original publication describing these mice (Tam et al., 2012, Dev Cell, PMID 22340501) did not report this information, and we were unable to obtain it directly from Genentech. Details regarding the genetic background of the Wld<sup>S</sup> and aPhr1 mutant mice are provided in their respective original publications, which are cited in our manuscript. Because the Gamage et al. study from the Deppmann laboratory did not report the sex or age of the animals used, we were unable to assess whether these variables might contribute to the differences observed between the two studies. Moreover, we are not aware of published evidence identifying sex or age as modifiers of structural axon preservation in axotomized peripheral nerve stumps in mouse models of delayed Wallerian degeneration. Furthermore, in the original publications describing the phenotypes of transgenic Nmnat2 and Wld<sup>S</sup> mice, as well as Sarm1 or Phr1 knockout mice, sex and age of the animals used in the Wallerian degeneration assays were not reported (PMIDs 23995269, 12106171, 22678360, 23665224). Although, to our knowledge, no large-scale systematic studies have been conducted, over the last 15 years we have never observed any sex-based differences in Wallerian degeneration phenotypes in these mutants exhibiting prominent axon protection. This topic was discussed informally at conferences, and we are also not aware of other investigators having observed such effects.
In response to the reviewer’s comment regarding “tone”, we made sure that our data and interpretations are presented in a professional, balanced, and objective manner, including a detailed discussion of potential alternative explanations for the discrepant findings.
(2) The DR6 knockout strain reported in Gamage et al. (2017) was on a C57BL/6.129S segregating background. Gamage et al. reported that loss of DR6 protected axons from Wallerian degeneration for up to 4 weeks, but importantly, only in 38.5% (5 out of 13) mice they examined. In the present paper, the authors speculate on possible causes for differences between the lack of effect seen here and the effects reported in Gamage et al., including possible spontaneous background mutations, epigenetic changes, genetic modifiers, neuroinflammation, and environmental differences. A likely explanation of the incomplete penetrance reported by Gamage et al. is the segregating genetic background and the presence of modifier loci between C57BL/6 and 129S. The authors do not report the genetic background of the mice used in this study, other than to note that the knockout strain was provided by the group in Gamage et al. However, if, for example, that mutation has been made congenic on C57BL/6 in the intervening years, this would be important to know. One could also argue that the results presented here are consistent with 8 out of 13 mice presented in Gamage et al.
(Response 2) As noted above, we now provide information on the genetic background of the mice in the revised manuscript, where available. We have not backcrossed the constitutive DR6 knockout mice obtained from the Deppmann laboratory (Gamage et al.) to a C57BL/6 background; our colony was maintained primarily through intercrosses of heterozygous animals. Similarly, the conditional DR6 mutant mice used in this study were also not backcrossed to C57BL/6 mice.
We respectfully hold a different view regarding the reviewer’s final point. We understand it is not appropriate to infer consistency between two datasets by disregarding the subset of results that do not align. By the same logic, it would be flawed to draw conclusions from the Gamage et al. study based solely on the single Wld<sup>S</sup> mouse out of five that did not show axon preservation after nerve injury. Selectively omitting conflicting data does not provide a valid basis for establishing phenotype concordance across studies.
To further strengthen our study, we note that we performed additional analyses on three more nerve samples from constitutive DR6 null mice during the revision process and have incorporated the resulting data in Fig. 1.
(3) Age is also an important variable. The protective effects of the spontaneous WldS mutation decrease with age, for example. It is unclear whether the possible protective effects of DR6 also change with age; perhaps this could explain the variable response seen in Gamage et al. and the lack of response seen here.
(Response 3) As discussed above, we now provide the age information for the mice used for the Wallerian degeneration assessments in the respective figure legends. To our knowledge, there are no prior reports suggesting that age is a significant determinant of structural axon preservation in the indicated mutants. Electrophysiological function and neuromuscular junction preservation decrease with age in axotomized Wld<sup>S</sup> mice (e.g., PMIDs 12231635, 19158292, 15654865), but these parameters are not subject of our study, and we have not studied them. Unfortunately, a direct comparison of ages between our DR6 mutant mice and those used in Gamage et al. (2017) is not possible, as the earlier study from the Deppmann laboratory did not report this information.
(4) It is unclear if sex is a factor, but this is part of why it should be reported.
(Response 4) We now report the requested sex information for our axon preservation analyses during nerve injury-induced Wallerian degeneration in the DR6 mouse models in Figs. 1 and 2.
(5) The authors also state that they do not see differences in the Schwann cell response to injury in the absence of DR6 that were reported in Gamage et al., but this is not an accurate comparison. In Gamage et al., they examined Schwann cells around axons that were protected from degeneration 2 and 4 weeks post-injury. Those axons had much thinner myelin, in contrast to axons protected by WldS or loss of Sarm1, where the myelin thickness remained relatively normal. Thus, Gamage et al. concluded that the protection of axons from degeneration and the preservation of Schwann cell myelin thickness are separate processes. Here, since no axon protection was seen, the same analysis cannot be done, and we can only say that when axons degenerate, the Schwann cells respond the same whether DR6 is expressed or not.
(Response 5) We appreciate the reviewer’s detailed comments. Our intention was not to directly compare our findings with those of Gamage et al. regarding the myelin behavior at these time points (because we never observed axon protection), but rather to note that we did not observe any DR6-dependent alterations in Schwann cell responses under conditions where axons undergo normal Wallerian degeneration. As the reviewer correctly points out, Gamage et al. analyzed Schwann cell myelin surrounding axons that were protected from degeneration for extended periods, a context fundamentally different from the complete lack of axon protection observed in our DR6-deficient models. Therefore, the specific dissociation between axon preservation and myelin maintenance claimed by Gamage et al. cannot be evaluated in our study. A statement to make this point clearer has been incorporated in the revised manuscript.
We fully agree with the reviewer’s concluding point: in our experiments, once axons degenerate, Schwann cell responses proceed similarly regardless of DR6 expression. This agreement reinforces one of the central conclusions of our work.
(6) The authors also take issue with Colombo et al. (2018), where it was reported that there is an increase in axon diameter and a change in the g-ratio (axon diameter to fiber diameter - the axon + myelin) in peripheral nerves in DR6 knockout mice. This change resulted in a small population of abnormally large axons that had thinner myelin than one would expect for their size. The change in g-ratio was specific to these axons and driven by the increased axon diameter, not decreased myelin thickness, although those two factors are normally loosely correlated. Here, the authors report no changes in axon size or g-ratio, but this could also be due to how the distribution of axon sizes was binned for analysis, and looking at individual data points in supplemental figure 3A, there are axons in the DR6 knockout mice that are larger than any axons in wild type. Thus, this discrepancy may be down to specifics and how statistics were performed or how histograms were binned, but it is unclear if the results presented here are dramatically at odds with the results in Colombo et al. (2018).
(Response 6) Several points raised by the reviewer appear to reflect differences in interpretation of the findings reported in Colombo et al. (2018). That study did not report altered myelination in DR6 null mice at stages when myelination is largely complete (P21). Instead, modest changes were observed at P1, which were reduced by P7, and P21 mutants were reported to be indistinguishable from controls. No analyses of peripheral nerves in older animals were presented, and the authors concluded in the discussion that myelination in young adult DR6 null mice appears normal. In contrast, our analysis of constitutive DR6 null mice at P1 does not reproduce the increase in the number of myelinated fibers per unit area reported by Colombo et al. We obtained similar results in the independent conditional DR6 knockout mouse line. Differences in nerve tissue processing, embedding, staining, or in the microscopic imaging and quantification of thinly myelinated axons in P1 sciatic nerve cross-sections may have contributed to the observed discrepancy. However, because the relevant methodological details were not described in Colombo et al., the underlying reasons for these differences cannot be determined and remain speculative.
(7) Finally, it is important to note that previously reported effects of DR6 inhibition, such as protection of cultured cortical neurons from beta-amyloid toxicity, are not necessarily the same as Wallerian degeneration of axons distal to an injury studied here. The negative results presented here, showing that loss of DR6 is not protective against Wallerian degeneration induced by injury, are important given the interest in DR6 as a therapeutic target, but they are specific to these mice and this mechanism of induced axon degeneration. The extent to which these findings contradict previous work is difficult to assess due to the lack of detail in describing the mouse experiments, and care should be taken in attempting to extrapolate these results to other disease contexts, such as ALS or Alzheimer's disease.
(Response 7) We agree with the reviewer’s point and emphasize that our manuscript carefully differentiates our data regarding the function of DR6 in Wallerian degeneration from the potential involvement of DR6 in other forms of axon degeneration. Our findings do not conflict with previous work on DR6 in the context of in vitro beta-amyloid and prion toxicity as well as in vitro models of ALS and multiple sclerosis. We believe these distinctions are explicitly and appropriately articulated throughout the entire manuscript and in more detail in the discussion section.
Reviewer #1 (Recommendations for the authors):
(1) The authors should include additional information about the mice used, including strain background for both the DR6 mice and the Cre transgenes crossed into the DR6 conditional knockout, the age of the mice when the nerve crush experiments were performed, and the sex distributions of the experimental cohorts. This information is critical for reproducibility in animal experiments, and that point is compounded here, where the major focus of this paper is taking issue with the reproducibility of previous work.
(Response 8) This information has been included in the revision. See above responses.
(2) In the abstract, reference 5 is cited as a study on the response to Schwann cells to injury in a DR6 background, but this probably should be reference 10.
(Response 9) This typo has been corrected.
(3) "Site-by-site comparison" in line 201 should be side-by-side?
(Response 10) This typo has been corrected.
(4) The paper contains a lot of self-evaluative wording, "surprising contrast," "compelling evidence," "robust results." Whether those adjectives apply should be for the reader to decide, and a drier, more objective tone in the presentation would improve the paper.
(Response 11) We agree that excessive self-evaluative wording can weaken objectivity. In the manuscript, such phrasing is used sparingly and intentionally to highlight differences from previously published studies, guide the reader, and convey scholarly judgment. We do not consider this limited use to be counterproductive. The adjectives “surprising,” “compelling,” and “robust” each appear only one to three times across the entire manuscript, and the specific phrase “robust results” does not appear at all.
(5) In Figure 2A, DR6-/-, there is no significant difference, but there is also a lot of variability, and one could argue the authors are seeing axon protection comparable to WldS in 40% of their samples (2/5), which is very similar to Gamage et al.
(Response 12) We respectfully disagree with this reasoning as it relies on selectively emphasizing only a subset of the data. Please also see our response #2 for more detailed discussion.
(6) Overall, the data presented here are convincing and support the conclusions drawn, but the paper needs to focus more on the negative results at hand and less on bashing previous studies, particularly when the results presented here do definitively show that the previous studies were incorrect and plausible explanations for differences in outcome exist.
(Response 13) We have carefully revisited the wording of the manuscript and are confident that our emphasis remains on the central negative finding that DR6 does not regulate axon degeneration and Schwann cell injury responses during Wallerian degeneration. We do not believe the manuscript “bashes” previous studies; nonetheless, we thoroughly re-examined all relevant sections to ensure that our language is neutral, accurate, and non-inflammatory. We believe the current phrasing presents our interpretations in an appropriately balanced, objective, and professional manner.
Reviewer #2 (Public review):
Summary:
This manuscript by Beirowski, Huang, and Babetto revisits the proposed role of Death Receptor 6 (DR6/Tnfrsf21) in Wallerian degeneration (WD). A prior study (Gamage et al., 2017) suggested that DR6 deletion delays axon degeneration and alters Schwann cell responses following peripheral nerve injury. Here, the authors comprehensively test this claim using two DR6 knockout mouse models (the line used in the earlier report plus a CMV-Cre derived floxed ko line) and multiple WD assays in vivo and in vitro, aligned with three positive controls, Sarm1 WldS and Phr1/Mycbp2 mutants. Contrary to the prior findings, they find no evidence that DR6 deletion affects axon degeneration kinetics or Schwann cell dynamics (assessed by cJun expression or [intact+degenerating] myelin abundance after injury) during WD. Importantly, in DRG explant assays, neurites from DR6-deficient mice degenerated at rates indistinguishable from controls. The authors conclude that DR6 is dispensable for WD, and that previously reported protective effects may have been due to confounding factors such as genetic background or spontaneous mutations.
Strengths:
The authors employ two independently generated DR6 knockout models, one overlapping with the previously published study, and confirm loss of DR6 expression by qPCR and Western blotting. Multiple complementary readouts of WD are applied (structural, ultrastructural, molecular, and functional), providing a robust test of the hypothesis.
Comparisons are drawn with established positive controls (WldS, SARM1, Phr1/Mycbp2 mutants), reinforcing the validity of the assays.
By directly addressing an influential but inconsistent prior report, the manuscript clarifies the role of DR6 and prevents potential misdirection of therapeutic strategies aimed at modulating WD in the PNS. The discussion thoughtfully considers possible explanations for the earlier results, including colony-specific second-site mutations that could explain the incomplete penetrance of the earlier reported phenotype of only 36%.
Weaknesses:
(1) The study focuses on peripheral nerves. The manuscript frequently refers to CNS studies to argue for consistency with their findings. It would be more accurate to frame PNS/CNS similarities as reminiscences rather than as consistencies (e.g., line 205ff in the Discussion).
(Response 14) Axon protection in all key genetic models of delayed axon degeneration, including Wld<sup>S</sup>, SARM1, Phr1/Mycbp2 mutants, has been demonstrated in both the peripheral and central nervous systems. This observation supports the view that core molecular mechanisms regulating axon degeneration are conserved across neuronal populations throughout the entire nervous system. We have scrutinized the wording in our manuscript and are not aware that we frequently refer to CNS studies in regards to axon degeneration. Nevertheless, we have replaced the term “consistent” to avoid potential ambiguity when we discuss the earlier study showing normal Wallerian degeneration in the optic nerves from DR6 knockout mice.
(2) The DRG explant assays are convincing, though the slight acceleration of degeneration in the DR6 floxed/Cre condition is intriguing (Figure 4E). Could the authors clarify whether this is statistically robust or biologically meaningful?
(Response 15) We thank the reviewer for noting this aspect of our in vitro data in Fig. 4. The difference observed in the DR6 floxed/Cre condition is statistically significant at the 6h time point following disconnection, as indicated by the p value shown in Fig. 4E. However, a similarly statistically significant acceleration of axon degeneration was not observed in DRG axotomy experiments using constitutive DR6 knockout preparations, although a trend toward more rapid axon breakdown is apparent at 6 h post-axotomy (Fig. 4B). These observations may suggest reduced stability of DR6-deficient axons in this specific neuron-only in vitro context. Further investigation would be required to determine the biological significance of this effect. In contrast, our in vitro quantitative analyses of the initiation and early phases of Wallerian degeneration (Fig. 2) revealed no evidence of accelerated axon disintegration in the DR6 mutant mouse models, highlighting potential differences between in vitro and in vitro systems.
(3) In the summary (line 43), the authors refer to Hu et al. (2013) (reference 5) as the study that previously reported AxD delay and SC response alteration after injury. However, this study did not investigate the PNS, and I believe the authors intended to reference Gamage et al. (2017) (reference 10) at this point.
(Response 16) Thanks for pointing this out. We have corrected this typo in the revised manuscript.
(4) In line 74ff of the results section, the authors claim that developmental myelination is not altered in DR6 mutants at postnatal day 1. However, the variability in Figure S2 appears substantial, and the group size seems underpowered to support this claim. Colombo et al. (2018) (reference 11) reported accelerated myelination at P1, but this study likewise appears underpowered. Possible reasons for these discrepancies and the large variability could be that only a defined cross-sectional area was quantified, rather than the entire nerve cross-section.
(Response 17) We confirm that the quantification of thinly myelinated axons was performed on entire sciatic nerves from P1 mouse pups, as described in the methods section in our original manuscript. The data shown in Fig. S2 were obtained from 5-9 pups per experimental group. Sample sizes were determined based on a priori power analyses using pilot data, which indicated that a minimum of five biological replicates was sufficient to detect statistically significant differences with acceptable confidence. Comparable sample sizes have been used in our previous studies and by other groups to assess early postnatal myelination (e.g., PMIDs 21949390, 28484008). Several published studies have reported analyses using 3-4 animals per group (e.g., PMIDs 28484008, 25310982, 29367382). For comparison, the study by Colombo et al. used 3-8 pups for the analysis presented in their Fig. 3. We note that the apparent variability in Fig. S2 may be accentuated by the scaling of the y-axis, which was chosen to ensure that individual data points are clearly resolved and visible.
(5) The authors stress the data of Gamage et al. (2017) on altered SC responses in DR6 mutants after injury. They employed cJun quantification to show that SC reprogramming after injury is not altered in DR6 mutants. This approach is valid and the conclusion trustworthy. Here, the addition of data showing the combined abundance of intact and degenerated myelin does not add much insight. However, Gamage et al. (2017) reported altered myelin thickness in a subset of axons at 14 days after injury, which is considerably later than the time points analyzed in the present study. While, in the Reviewer's view, the thin myelin observed by Gamage et al. in fact resembles remyelination, the authors may wish to highlight the difference in the time points analyzed.
(Response 18) We consider the additional quantification of the area occupied by intact myelin and myelin debris to provide complementary information that supports the c-Jun-based conclusion that Schwann cell injury responses are normal in DR6-deficient nerves following lesion. We agree with this reviewer that the thin myelin observed by Gamage et al. resembles remyelination, raising the possibility that axon regeneration occurred into the distal nerve stump at the studied 14d post-injury time point (see their Fig. 3). This may have been interpreted as axon protection in this study. In our study, it was impossible to examine such myelin effects since axon protection was never observed in any of the DR6 mutant models at any of the time point we investigated. We have incorporated appropriate additional text to highlight this difference. See also response #5 above.
Reviewer #3 (Public review):
Summary:
The authors revisit the role of DR6 in axon degeneration following physical injury (Wallerian degeneration), examining both its effects on axons and its role in regulating the Schwann cell response to injury. Surprisingly, and in contrast to previous studies, they find that DR6 deletion does not delay the rate of axon degeneration after injury, suggesting that DR6 is not a mediator of this process.
Overall, this is a valuable study. As the authors note, the current literature on DR6 is inconsistent, and these results provide useful new data and clarification. This work will help other researchers interpret their own data and re-evaluate studies related to DR6 and axon degeneration.
Strengths:
(1) The use of two independent DR6 knockout mouse models strengthens the conclusions, particularly when reporting the absence of a phenotype.
(2) The focus on early time points after injury addresses a key limitation of previous studies. This approach reduces the risk of missing subtle protective phenotypes and avoids confounding results with regenerating axons at later time points after axotomy.
Weaknesses:
(1) The study would benefit from including an additional experimental paradigm in which DR6 deficiency is expected to have a protective effect, to increase confidence in the experimental models, and to better contextualize the findings within different pathways of axon degeneration. For example, DR6 deletion has been shown in more than one study to be partially axon protective in the NGF deprivation model in DRGs in vitro. Incorporating such an experiment could be straightforward and would strengthen the paper, especially if some of the neuroprotective effects previously reported are confirmed.
(Response 19) We thank the reviewer for these suggestions. We would like to highlight that our study addresses the role of DR6 in Wallerian degeneration, whereas in vitro NGF deprivation has been used to model developmental axon pruning. Previous work indicates fundamental biological differences between these regressive pathways regulating the stereotyped removal of axon segments. We feel that studying this alternative form of axon degeneration is beyond the scope of the current work and could be addressed in a separate manuscript. Although additional tests will be needed, we note that our preliminary data using samples from both DR6 knockout mouse models suggest no axon protection after NGF-deprivation in DRG neuron preparations in our hands (deprivation of the growth factor and administration of anti-NGF antibody).
(2) The quality of some figures could be improved, particularly the EM images in Figure 2. As presented, they make it difficult to discern subtle differences.
(Response 20) We have pseudocolored intact (turquoise) and degenerated (magenta) myelinated fibers on the high-resolution semithin micrographs (not electron micrographs) in the new Fig. 2 to make the distinction between the two fiber categories clearer.
Reviewer #3 (Recommendations for the authors):
(1) Line 121: The authors mention toluidine blue staining, but it does not appear to be shown in Figure S5.
(Response 21) This appears to be a misunderstanding. Fig. S5A shows the ultrastructure of dedifferentiated Schwann cells in transmission electron micrographs, while Figs. S5B and C show quantification of the area occupied by myelin sheaths and myelin debris profiles on osmium tetroxide and toluidine blue stained nerve sections from the two DR6 mutant models, based on semithin light microscopy. These are two different aspects of the analysis. The text has been modified in the revised manuscript to make the distinction clearer.
(2) Line 175: The authors should add NMNAT2 to the list of enzymes implicated in the regulation of Wallerian degeneration in mammals.
(Response 22) Nmnat2 and a literature reference (Milde et al., 2013) has been incorporated in the discussion of the revised manuscript to address this point.
(3) Line 201: Please correct the typo "site-by-site" to "side-by-side."
(Response 23) This typo has been corrected.
eLife Assessment
This fundamental work significantly advances our understanding of how contact-dependent antagonism enables keystone bacteria to establish and maintain their niche over time. The evidence obtained is convincing, supporting most of the conclusions drawn. This work will be of significant interest to the microbiome research community.
Reviewer #1 (Public review):
Summary:
In this study, the authors investigate the physiological role of the Type VI secretion system (T6SS) in a naturally evolved gut microbiome derived from wild mice (the WildR microbiome). Focusing on Bacteroides acidifaciens, the authors use newly developed genetic tools and strain-replacement strategies to test how T6SS-mediated antagonism influences colonization, persistence, and fitness within a complex gut community. They further show that the T6SS resides on an integrative and conjugative element (ICE), is distributed among select community members, and can be horizontally transferred, with context-dependent effects on colonization and persistence. The authors conclude that the T6SS stabilizes strain presence in the gut microbiome while imposing ecological and physiological constraints that shape its value across contexts.
This study is likely to have a significant impact on the microbiome field by moving experimental tests of T6SS function out of simplified systems and into a naturally co-evolved gut community. The WildR system, together with the strain replacement strategy, ICE-seq approach, and genetic toolkit, represents a powerful and reusable platform for future mechanistic studies of microbial antagonism and mobile genetic elements in vivo.
The datasets, including isolate genomes, metagenomes, and ICE distribution maps, will be a valuable community resource, particularly for researchers interested in strain-resolved dynamics, horizontal gene transfer, and ecological context dependence. Even where mechanistic resolution is incomplete, the work provides a strong experimental foundation upon which such questions can be directly addressed.
Overall, this study occupies a space between system building and mechanistic dissection. The authors demonstrate that the T6SS influences persistence and community structure in vivo, but the physiological basis of these effects remains unresolved. Interpreting the results as evidence of fitness costs or selective advantage, therefore, requires caution, as multiple ecological and host-mediated processes could produce similar abundance trajectories.
Placing the findings within the broader literature on microbial antagonism, particularly work emphasizing measurable costs, benefits, and tradeoffs, would help readers better contextualize what is directly demonstrated here versus what remains an open question. Viewed in this light, the principal contribution of the study is to show that such questions can now be addressed experimentally in a realistic gut ecosystem.
Strengths:
A major strength of this study is that it directly interrogates the physiological role of the T6SS in a naturally evolved gut microbiome, rather than relying on simplified pairwise or in vitro systems. By working within the WildR community, the authors advance beyond descriptive surveys of T6SS prevalence and address function in an ecologically relevant context.
The authors provide clear genetic evidence that Bacteroides acidifaciens uses a T6SS to antagonize co-resident Bacteroidales, and that loss of T6SS function specifically compromises long-term persistence without affecting initial colonization. This temporal separation is well designed and supports the conclusion that the T6SS contributes to maintenance rather than establishment within the community.
Another strength is the identification of the T6SS on an integrative and conjugative element (ICE) and the demonstration that this element is distributed among, and exchanged between, community members. The use of ICE-seq to track distribution and transfer provides strong support for horizontal mobility and adds mechanistic depth to the study.
Finally, the transfer of the T6SS-ICE into Phocaeicola vulgatus and the observation of context-dependent colonization benefits followed by decline is a compelling result that moves the study beyond simple "T6SS is beneficial" narratives and highlights ecological contingency.
Weaknesses:
Despite these strengths, there is a mismatch between the precision of the claims and the precision of the measurements, particularly regarding fitness costs, physiological burden, and the mechanistic role of the T6SS.
First, while the authors conclude that the T6SS "stabilizes strain presence" and that its value is constrained by fitness costs, these costs are not directly measured. Persistence, abundance trajectories, and eventual loss are informative outcomes, but they do not uniquely identify fitness tradeoffs. Decline could arise from multiple non-exclusive mechanisms, including community restructuring, host-mediated effects, incompatibilities of the ICE in new hosts, or ecological retaliation, none of which are disentangled here.
Second, the manuscript frames the T6SS as having a defined physiological role, yet the data do not resolve which physiological processes are under selection. The experiments demonstrate that T6SS activity affects persistence, but they do not distinguish whether this occurs via direct killing, resource release, niche modification, or higher-order community effects. As a result, "physiological role" remains underspecified and risks being conflated with ecological outcome.
Third, although the authors emphasize context dependence, the study offers limited quantitative insight into what aspects of context matter. Differences between native and recipient hosts, or between early and late colonization phases, are described but not mechanistically interrogated, making it difficult to generalize beyond the specific cases examined.
Fourth is the lack of engagement with recent experimental literature demonstrating functional roles of the T6SS beyond simple interference competition. While the authors focus on persistence and competitive outcomes, they do not adequately situate their findings within recent work demonstrating that T6SS-mediated antagonism can serve additional physiological functions, including resource acquisition and DNA uptake, thereby linking killing to measurable benefits and tradeoffs. The absence of this literature makes it difficult to place the authors' conclusions about physiological role and fitness cost within the current conceptual framework of the field. Without this context, the physiological interpretation of the results remains incomplete, and alternative functional explanations for the observed dynamics are underexplored.
A further limitation concerns the taxonomic scope of the functional analysis. The authors state that the role of the T6SS in the murine environment is functionally investigated using genetically tractable Bacteroides species, citing the lack of genetic tools for Mucispirillum schaedleri. While this is a reasonable, practical choice, it means that a substantial fraction of T6SS-encoding species in the WildR community are not experimentally interrogated. Consequently, conclusions about the role of the T6SS in the murine gut necessarily reflect the subset of taxa that are genetically accessible and may not fully capture community-level or niche-specific functions of T6SS activity. Given that M. schaedleri is represented as a metagenome-assembled genome, its isolation and genetic manipulation would be technically challenging. Nonetheless, explicitly acknowledging this limitation and slightly tempering claims of generality would strengthen the manuscript.
Finally, several interpretations would benefit from more cautious language. In particular, claims invoking fitness costs, selective advantage, or physiological burden should be explicitly framed as inferences from persistence dynamics, rather than as direct measurements, unless supported by additional quantitative fitness or growth assays.
Reviewer #2 (Public review):
Summary:
In this study, the authors set out to determine how a contact-dependent bacterial antagonistic system contributes to the ability of specific bacterial strains to persist within a complex, native gut community derived from wild animals. Rather than focusing on simplified or artificial models, the authors aimed to examine this system in a biologically realistic setting that captures the ecological complexity of the gut environment. To achieve this, they combined controlled laboratory experiments with animal colonization studies and sequencing-based tracking approaches that allow individual strains and mobile genetic elements to be followed over time.
Strengths:
A major strength of the work is the integration of multiple complementary approaches to address the same biological question. The use of defined but complex communities, together with in vivo experiments, provides a strong ecological context for interpreting the results. The data consistently show that the antagonistic system is not required for initial establishment but plays a critical role in long-term strain persistence. This insight that moves beyond traditional invasion-based views of microbial competition. The observation that transferable genetic elements can confer only temporary advantages, and may impose longer-term costs depending on community context, adds important nuance to current understanding of microbial fitness.
Weaknesses:
Overall, there is not a lack of evidence, but a deliberate trade-off between ecological realism and mechanistic resolution, which leaves some causal pathways open to interpretation.
Reviewer #3 (Public review):
Summary:
Shen et al. investigate the contribution of the type VI secretion system of Bacteroidales in the gut microbiome assembly and targeting of closely related species. They demonstrate that B. acidifaciens relies on T6SS-mediated antagonism to prevent displacement by co-resident Bacteroidales and other members of the microbiome, allowing B. acidifaciens to persist in the gut.
Strengths:
Using a gnotobiotic model colonized with a wild-mouse microbiome is a significant strength of this study. This approach allows tracking of microbiome changes over time and directly examining targeting by Bacteroidales carrying T6SS in a more natural setting. The development of ICE-seq for mapping the distribution of the T6SS in the microbiome is remarkable, enabling the study of how this bacterial weapon is transferred between microbiome members without requiring long-read metagenomics methods.
Weaknesses:
Some conclusions are based on only four mice per condition. The author should consider increasing the sample size.
Overall, the authors successfully achieved their objectives, and their experimental design and results support their findings. As mentioned in the discussion, it would be important to investigate the role of the T6SS in resilience to disturbances in the microbiome, such as antibiotics, diet, or pathogen invasion. This work represents a step forward in understanding how contact-dependent competition influences the gut microbiome in relevant ecological contexts.
Author response:
We appreciate that the reviewers provided an overall positive assessment of our manuscript and offered constructive suggestions for improvement. All three reviewers noted that a key strength of our study is the implementation of a gut microbiome model for the characterization of interbacterial antagonism pathways such as the type VI secretion system (T6SS) that approaches natural complexity. They note our work represents a significant advance in microbiome research, and generates resources that will be of use to many researchers in the field. Two of the reviewers point out that the complexity of our model limits the nature of measurements we can make, and suggest we temper the strength of the some of the conclusions we draw. As noted in more detail below, in our revised manuscript, we will be more precise in the wording we use to characterize our findings, and we will be more explicit about what the measurements we are able to make allow us to conclude about the physiological role of the T6SS in the gut microbiome.
Reviewer #1 (Public review):
Summary:
In this study, the authors investigate the physiological role of the Type VI secretion system (T6SS) in a naturally evolved gut microbiome derived from wild mice (the WildR microbiome). Focusing on Bacteroides acidifaciens, the authors use newly developed genetic tools and strain-replacement strategies to test how T6SS-mediated antagonism influences colonization, persistence, and fitness within a complex gut community. They further show that the T6SS resides on an integrative and conjugative element (ICE), is distributed among select community members, and can be horizontally transferred, with context-dependent effects on colonization and persistence. The authors conclude that the T6SS stabilizes strain presence in the gut microbiome while imposing ecological and physiological constraints that shape its value across contexts.
This study is likely to have a significant impact on the microbiome field by moving experimental tests of T6SS function out of simplified systems and into a naturally co-evolved gut community. The WildR system, together with the strain replacement strategy, ICE-seq approach, and genetic toolkit, represents a powerful and reusable platform for future mechanistic studies of microbial antagonism and mobile genetic elements in vivo.
The datasets, including isolate genomes, metagenomes, and ICE distribution maps, will be a valuable community resource, particularly for researchers interested in strain-resolved dynamics, horizontal gene transfer, and ecological context dependence. Even where mechanistic resolution is incomplete, the work provides a strong experimental foundation upon which such questions can be directly addressed.
Overall, this study occupies a space between system building and mechanistic dissection. The authors demonstrate that the T6SS influences persistence and community structure in vivo, but the physiological basis of these effects remains unresolved. Interpreting the results as evidence of fitness costs or selective advantage, therefore, requires caution, as multiple ecological and host-mediated processes could produce similar abundance trajectories.
Placing the findings within the broader literature on microbial antagonism, particularly work emphasizing measurable costs, benefits, and tradeoffs, would help readers better contextualize what is directly demonstrated here versus what remains an open question. Viewed in this light, the principal contribution of the study is to show that such questions can now be addressed experimentally in a realistic gut ecosystem.
We thank the reviewer for this thoughtful summary of our study. We were glad to read they conclude our work will have a significant impact on the microbiome field and that the resources we have developed will be of value to the community.
Strengths:
A major strength of this study is that it directly interrogates the physiological role of the T6SS in a naturally evolved gut microbiome, rather than relying on simplified pairwise or in vitro systems. By working within the WildR community, the authors advance beyond descriptive surveys of T6SS prevalence and address function in an ecologically relevant context.
The authors provide clear genetic evidence that Bacteroides acidifaciens uses a T6SS to antagonize co-resident Bacteroidales, and that loss of T6SS function specifically compromises long-term persistence without affecting initial colonization. This temporal separation is well designed and supports the conclusion that the T6SS contributes to maintenance rather than establishment within the community.
Another strength is the identification of the T6SS on an integrative and conjugative element (ICE) and the demonstration that this element is distributed among, and exchanged between, community members. The use of ICE-seq to track distribution and transfer provides strong support for horizontal mobility and adds mechanistic depth to the study.
Finally, the transfer of the T6SS-ICE into Phocaeicola vulgatus and the observation of context-dependent colonization benefits followed by decline is a compelling result that moves the study beyond simple "T6SS is beneficial" narratives and highlights ecological contingency.
We appreciate this detailed and nuanced characterization of the strengths of our study.
Weaknesses:
Despite these strengths, there is a mismatch between the precision of the claims and the precision of the measurements, particularly regarding fitness costs, physiological burden, and the mechanistic role of the T6SS.
We acknowledge that in some places, our manuscript could benefit from greater precision in the language we use when linking the outcomes we observe in our study to their potential underlying causes. Specific revisions we propose to address this concern are described below.
First, while the authors conclude that the T6SS "stabilizes strain presence" and that its value is constrained by fitness costs, these costs are not directly measured. Persistence, abundance trajectories, and eventual loss are informative outcomes, but they do not uniquely identify fitness tradeoffs. Decline could arise from multiple non-exclusive mechanisms, including community restructuring, host-mediated effects, incompatibilities of the ICE in new hosts, or ecological retaliation, none of which are disentangled here.
We agree that multiple mechanisms could explain why P. vulgatus carrying a T6SS-encoding ICE declines over time. Our use of the term “fitness cost” to describe this trend was not meant to imply any particular underlying mechanism, but was rather our attempt to characterize the phenotypic outcome we observed in simplified terms. We note that ecological context is an important determinant of the fitness cost or benefit of any given trait, and our study sheds light on the importance of the presence of the WildR community and the mouse intestinal environment to the fitness contribution of the ICE to P. vulgatus. Nonetheless, to avoid implying an overly simplistic interpretation of our results, we propose to modify the language used in the manuscript when describing the contribution of the T6SS to species persistence in WildR-colonized mice.
Second, the manuscript frames the T6SS as having a defined physiological role, yet the data do not resolve which physiological processes are under selection. The experiments demonstrate that T6SS activity affects persistence, but they do not distinguish whether this occurs via direct killing, resource release, niche modification, or higher-order community effects. As a result, "physiological role" remains underspecified and risks being conflated with ecological outcome.
We acknowledge that our study does not fully resolve the physiological processes under selection that mediate role of the T6SS in maintaining B. acidifaciens populations in WildR-colonized mice. Indeed, several of the outcomes of T6SS activity the reviewer lists, such as target cell killing and nutrient release, are inextricably linked and thus inherently difficult to disentangle. We note that we did attempt to measure higher-order community effects of T6SS activity with metagenomic sequencing, but acknowledge that this approach may not have been sufficiently sensitive to detect small community shifts mediated by a relatively low-abundance species. To address the concern that our current framing implies more of a mechanistic understanding that our study achieves, we propose to substitute “ecological” for “physiological” where appropriate when summarizing our key findings.
Third, although the authors emphasize context dependence, the study offers limited quantitative insight into what aspects of context matter. Differences between native and recipient hosts, or between early and late colonization phases, are described but not mechanistically interrogated, making it difficult to generalize beyond the specific cases examined.
We are not entirely clear what the reviewer means by “differences between native and recipient hosts”, but we agree that additional quantitative studies will be needed to address the generalizability of our findings. Future studies are also needed to address the mechanistic basis for the difference in the benefit conferred by the T6SS that we observed between P. vulgatus and B. acidifaciens.
Fourth is the lack of engagement with recent experimental literature demonstrating functional roles of the T6SS beyond simple interference competition. While the authors focus on persistence and competitive outcomes, they do not adequately situate their findings within recent work demonstrating that T6SS-mediated antagonism can serve additional physiological functions, including resource acquisition and DNA uptake, thereby linking killing to measurable benefits and tradeoffs. The absence of this literature makes it difficult to place the authors' conclusions about physiological role and fitness cost within the current conceptual framework of the field. Without this context, the physiological interpretation of the results remains incomplete, and alternative functional explanations for the observed dynamics are underexplored.
We thank the reviewer for specifically highlighting the potential pertinence of this literature to our study. Indeed, we did not cite studies indicating a link between T6SS activity and the uptake of DNA and other resources released by targeted cells. As we note above, the release of intracellular contents from target cells is an inevitable consequence of the delivery of lytic effectors. Thus, distinguishing between fitness benefits conferred from the elimination of competitor species and those arising from scavenging the nutrients released during this process is not straightforward. Measuring the benefits deriving from the uptake of certain released molecules, such as DNA, was not immediately feasible in the system employed in this study and instead we focused on the direct lytic consequences of the effectors delivered via the T6SS. We will revise our Discussion to include reference to how downstream consequences of T6SS activity on target cells could impact the community, and thus the adaptive role of the T6SS in the microbiome.
A further limitation concerns the taxonomic scope of the functional analysis. The authors state that the role of the T6SS in the murine environment is functionally investigated using genetically tractable Bacteroides species, citing the lack of genetic tools for Mucispirillum schaedleri. While this is a reasonable, practical choice, it means that a substantial fraction of T6SS-encoding species in the WildR community are not experimentally interrogated. Consequently, conclusions about the role of the T6SS in the murine gut necessarily reflect the subset of taxa that are genetically accessible and may not fully capture community-level or niche-specific functions of T6SS activity. Given that M. schaedleri is represented as a metagenome-assembled genome, its isolation and genetic manipulation would be technically challenging. Nonetheless, explicitly acknowledging this limitation and slightly tempering claims of generality would strengthen the manuscript.
The reviewer points out that studying the T6SS activity in M. schadleri would potentially expand the generality of our claims. We agree that having an isolate of this species along with genetic tools for its manipulation would allow us to probe the importance of the T6SS in the gut microbiome more broadly. At the suggestion of the reviewer, we will add explicit mention for the need to develop such tools, an endeavor that lies outside of the scope of the current study.
Finally, several interpretations would benefit from more cautious language. In particular, claims invoking fitness costs, selective advantage, or physiological burden should be explicitly framed as inferences from persistence dynamics, rather than as direct measurements, unless supported by additional quantitative fitness or growth assays.
We agree with the reviewer that invoking fitness costs, selective advantages or physiological burdens should be done cautiously, and in our revised manuscript we will carefully re-evalute our usage of those terms. However, we would also argue invoking fitness costs and benefits when describe strain persistence dynamics in mice has substantial precedent in the literature ((Feng et al. 2020, Brown et al. 2021, Park et al. 2022, Segura Munoz et al. 2022), to list a handful of representative examples published by different groups). It is unclear to us what additional in vivo growth measurements could be taken to substantiate our claim that the T6SS provides a fitness benefit to B. acidifaciens during prolonged gut colonization, or that carrying the ICE imposes a fitness cost on P. vulgatus during long-term colonization. Our in vitro experiments evaluating the competitiveness conferred by T6SS activity provide a measure of insight into its fitness benefits, but as our in vivo strain persistence data and the work of many others show, in vitro measurements do not necessarily capture in vivo parameters.
Reviewer #2 (Public review):
Summary:
In this study, the authors set out to determine how a contact-dependent bacterial antagonistic system contributes to the ability of specific bacterial strains to persist within a complex, native gut community derived from wild animals. Rather than focusing on simplified or artificial models, the authors aimed to examine this system in a biologically realistic setting that captures the ecological complexity of the gut environment. To achieve this, they combined controlled laboratory experiments with animal colonization studies and sequencing-based tracking approaches that allow individual strains and mobile genetic elements to be followed over time.
Strengths:
A major strength of the work is the integration of multiple complementary approaches to address the same biological question. The use of defined but complex communities, together with in vivo experiments, provides a strong ecological context for interpreting the results. The data consistently show that the antagonistic system is not required for initial establishment but plays a critical role in long-term strain persistence. This insight that moves beyond traditional invasion-based views of microbial competition. The observation that transferable genetic elements can confer only temporary advantages, and may impose longer-term costs depending on community context, adds important nuance to current understanding of microbial fitness.
We thank the reviewer for the positive feedback and are glad they agree our study provides new insight into the role of interbacterial antagonism in natural communities.
Weaknesses:
Overall, there is not a lack of evidence, but a deliberate trade-off between ecological realism and mechanistic resolution, which leaves some causal pathways open to interpretation.
The reviewer makes a good point that the complexity of the experimental system we employ precludes some lines of experimentation that would yield more mechanistic information. As the reviewer notes, we were aware of the tradeoff between mechanistic resolution and ecological realism when selecting our experimental system. Our deliberate choice to favor biological complexity over mechanistic clarity in this study stemmed from our perception that a major gap in understanding of the T6SS and other antagonism pathways lies in defining their ecological function in complex microbial communities.
Reviewer #3 (Public review):
Summary:
Shen et al. investigate the contribution of the type VI secretion system of Bacteroidales in the gut microbiome assembly and targeting of closely related species. They demonstrate that B. acidifaciens relies on T6SS-mediated antagonism to prevent displacement by co-resident Bacteroidales and other members of the microbiome, allowing B. acidifaciens to persist in the gut.
Strengths:
Using a gnotobiotic model colonized with a wild-mouse microbiome is a significant strength of this study. This approach allows tracking of microbiome changes over time and directly examining targeting by Bacteroidales carrying T6SS in a more natural setting. The development of ICE-seq for mapping the distribution of the T6SS in the microbiome is remarkable, enabling the study of how this bacterial weapon is transferred between microbiome members without requiring long-read metagenomics methods.
We thank the reviewer for their enthusiasm toward our study.
Weaknesses:
Some conclusions are based on only four mice per condition. The author should consider increasing the sample size.
We agree that in some experiments it would be beneficial to increase the sample size from four mice. However, the experiments we performed for this study are time and resource intensive. Additionally, the experiments on which we base our primary conclusions were all independently replicated with similar results. Given these factors, we determined that the extra confidence that might be afforded by increasing our sample size did not merit the delay in publication and investment in resources that would be required.
Overall, the authors successfully achieved their objectives, and their experimental design and results support their findings. As mentioned in the discussion, it would be important to investigate the role of the T6SS in resilience to disturbances in the microbiome, such as antibiotics, diet, or pathogen invasion. This work represents a step forward in understanding how contact-dependent competition influences the gut microbiome in relevant ecological contexts.
We agree that investigating the role of the T6SS during perturbations of the microbiome is a key next step for this work and thank the reviewer for highlighting this important future direction.
References
Brown, E. M., H. Arellano-Santoyo, E. R. Temple, Z. A. Costliow, M. Pichaud, A. B. Hall, K. Liu, M. A. Durney, X. Gu, D. R. Plichta, C. A. Clish, J. A. Porter, H. Vlamakis and R. J. Xavier (2021). "Gut microbiome ADP-ribosyltransferases are widespread phage-encoded fitness factors." Cell Host Microbe 29(9): 1351-1365 e1311.
Feng, L., A. S. Raman, M. C. Hibberd, J. Cheng, N. W. Griffin, Y. Peng, S. A. Leyn, D. A. Rodionov, A. L. Osterman and J. I. Gordon (2020). "Identifying determinants of bacterial fitness in a model of human gut microbial succession." Proc Natl Acad Sci U S A 117(5): 2622-2633.
Park, S. Y., C. Rao, K. Z. Coyte, G. A. Kuziel, Y. Zhang, W. Huang, E. A. Franzosa, J. K. Weng, C. Huttenhower and S. Rakoff-Nahoum (2022). "Strain-level fitness in the gut microbiome is an emergent property of glycans and a single metabolite." Cell 185(3): 513-529 e521.
Segura Munoz, R. R., S. Mantz, I. Martinez, F. Li, R. J. Schmaltz, N. A. Pudlo, K. Urs, E. C. Martens, J. Walter and A. E. Ramer-Tait (2022). "Experimental evaluation of ecological principles to understand and modulate the outcome of bacterial strain competition in gut microbiomes." ISME J 16(6): 1594-1604.
Guide de Scolarisation des Élèves Présentant des Troubles à Expression Comportementale
Ce document synthétise les stratégies et outils destinés aux enseignants pour scolariser efficacement les élèves manifestant des troubles du comportement.
La distinction fondamentale repose sur la différence entre une opposition ponctuelle (réactionnelle et passagère) et des troubles du comportement avérés (crises intenses, incapacité de régulation, dangerosité).
La prise en charge repose sur trois piliers :
1. La Prévention : Création d’un environnement sécurisant par une organisation spatiale et temporelle stable et une posture d'enseignant prévisible.
2. L’Adaptation : Utilisation d’outils de structuration (contrats de comportement, thermomètres émotionnels, espaces de répit) pour répondre aux besoins spécifiques de l'élève.
3. La Gestion de Crise : Application de protocoles de désescalade et mise en sécurité, suivies d'une phase d'analyse rigoureuse pour ajuster les interventions futures.
L'objectif central est de passer d'une gestion réactive à une approche proactive, visant l'apaisement de l'élève et la préservation du climat d'apprentissage pour l'ensemble de la classe.
--------------------------------------------------------------------------------
Il est crucial pour l'enseignant de diagnostiquer la nature de la perturbation afin d'y apporter la réponse appropriée.
Le tableau suivant distingue les deux types de manifestations comportementales :
| Caractéristiques | Opposition Ponctuelle | Troubles du Comportement Avérés | | --- | --- | --- | | Manifestations | Refus temporaire, frustration verbale, énervement bref. | Crises fréquentes, violences physiques (soi, autres, matériel), agressivité constante. | | Capacité de régulation | Retrouve son calme après un rappel ou une redirection. | Incapacité à se réguler seul, même avec soutien. | | Origine | Fatigue, difficulté de compréhension, test des limites. | Épuisement émotionnel ou sensoriel, déconnecté de la situation immédiate. | | Impact | Ne perturbe pas durablement la classe. | Perturbation majeure du climat de classe et des apprentissages. |
Une crise majeure se définit par une perte totale de contrôle. Elle est caractérisée par une intensité forte (hurlements, violences), une durée significative (minutes à heures), et un danger potentiel. Dans cet état, l'élève n'est plus dans une logique de calcul ou d'opposition délibérée.
--------------------------------------------------------------------------------
La prévention consiste à être proactif pour minimiser les déclencheurs comportementaux.
• Stabilité Spatiale : Les places doivent être fixées. L'enseignant doit voir et être vu de tous. Les déplacements doivent être aisés et les procédures de rangement enseignées.
• Stabilité Temporelle : Utilisation d'un emploi du temps hebdomadaire stable, affichage de l'emploi du temps quotidien et mise en place de rituels et routines systématiques.
L'enseignant doit incarner un modèle de stabilité :
• Élaborer le règlement de classe avec les élèves et l'afficher.
• Formuler les règles de manière affirmative (expliciter le comportement attendu plutôt que l'interdit).
• Avoir des réactions prévisibles et mesurées.
• Agir avec crédibilité : "Dire ce que je fais et faire ce que je dis."
--------------------------------------------------------------------------------
Chaque besoin identifié doit correspondre à un aménagement technique ou pédagogique précis.
• Espace : Prévoir un espace de travail individualisé et un espace d'apaisement (coin détente avec livres, casque de musique) dont le temps d'accès est limité par un timer.
• Temps : Utiliser des supports visuels (horloges, sabliers, timers) et des emplois du temps individualisés pour rendre les durées concrètes.
• Émotions : Utiliser le "thermomètre des émotions" ou "l'humeur du jour" pour aider l'élève à identifier son état interne.
• Relation aux autres : Mettre en place des signaux discrets, comme le Tétra-aide, pour que l'élève puisse appeler à l'aide sans perturber le groupe.
Cet outil d'engagement mutuel vise à valoriser les comportements adaptés :
• Fixation d'objectifs simples.
• Auto-évaluation quotidienne par l'élève.
• Valorisation systématique des réussites (parole positive ou accès à une activité appréciée).
• Implication de la famille dans le suivi des progrès.
Il est nécessaire d'adapter les exigences aux capacités de l'élève (via PAP ou PPRE) :
• Détailler spécifiquement le comportement attendu pour chaque tâche.
• Privilégier les appels positifs ("Tu rejoins la table") plutôt que les questions ouvertes.
• Ressources numériques : Utiliser des sites comme Cap Ecole Inclusive ou Araasac (pictogrammes), et des logiciels comme Lire Couleur (aide à la lecture) ou Dicom (prédiction de mots).
--------------------------------------------------------------------------------
La gestion d'une crise suit un cycle spécifique nécessitant des interventions ciblées à chaque phase.
L'objectif est d'intervenir idéalement dès la phase d'activation pour éviter l'escalade :
1. Calme
2. Activation : Signes subtils (anxiété, erreurs de jugement, maux de tête/ventre).
3. Agitation / Accélération : Difficulté à réguler la parole, besoin d'attention, agitation psychomotrice.
4. Point culminant (Crise) : Perte de contrôle.
5. Décélération / Récupération
• Fermeté : Sur les actes inacceptables (violence, jet de matériel) entraînant un écart immédiat du groupe.
• Apaisement : Utiliser une voix basse et des paroles contenantes ("Tout va bien", "Je vais t'aider", "Ton bien-être compte pour moi").
• Protocole : Un protocole écrit doit définir qui prend en charge l'élève, qui gère le reste de la classe, et qui prévient la famille ou les secours (le 15 en cas de gravité extrême).
Une fois le calme revenu, un travail d'analyse est indispensable :
• Constater : Consigner les faits (avant, pendant, après).
• Analyser : Échanger avec l'élève et la famille pour identifier les déclencheurs ou les éléments renforçateurs.
• Réajuster : Proposer de nouvelles adaptations ou modifier le protocole de crise si nécessaire.
--------------------------------------------------------------------------------
"Dans une crise majeure, l’élève n’est pas dans une logique d’opposition ou de calcul, mais dans un état d’épuisement émotionnel ou sensoriel."
"Dire ce que je fais et faire ce que je dis. (paroles suivies des actions)"
"L’objectif de ce protocole est de viser l’extinction des crises en gardant les exigences pour l’élève."
hello friends
§ 103. Recall Any elected official or official appointed to elective office holding a Town office may be recalled by the following process: (1) A petition signed by not less than 35 percent of the number of registered voters voting in the last annual Town meeting shall be filed with the Town Clerk. (2) The petition shall request a special Town meeting for the purpose of recalling a specific person or persons and shall state the office that they hold. (3) The Selectboard shall act upon the petition within seven days by warning a special Town meeting. (4) The Town meeting shall be held not less than 30 days and not more than 40 days from the date of the receipt of the petition. (5) The special Town meeting shall be warned as provided by the general law of this State. (6) At least four days but not more than 15 days prior to the voting, an informational meeting shall be held at which time both the elected official and the proponents for the recall shall have the opportunity to discuss the issue of recall. The informational meeting shall be warned in conjunction with the warning for the special Town meeting. (7) Voting on the issue of recall shall be by Australian ballot. A majority of the legal votes cast shall be needed to effect a recall of that official.
La Bienveillance en Milieu Scolaire : Enjeux, Défis et Pratiques Professionnelles
Ce document synthétise les échanges issus d'une table ronde portant sur le concept de bienveillance à l'école.
Loin d'être synonyme de laxisme ou de complaisance, la bienveillance est définie comme une condition essentielle de l'équité et de l'efficacité du système éducatif, particulièrement pour les élèves les plus vulnérables.
Elle repose sur une tension constructive entre exigence et soutien, visant le développement à long terme de l'élève.
Sa mise en œuvre nécessite une clarification conceptuelle pour lever les résistances professionnelles, l'adoption de gestes professionnels spécifiques (feedback positif, écoute active) et une réinvention des espaces et des modalités d'évaluation.
En somme, la bienveillance est un levier de réussite qui engage tant la posture individuelle de l'enseignant que la stratégie collective de l'établissement.
--------------------------------------------------------------------------------
La bienveillance en éducation souffre souvent de représentations simplistes ou erronées. Il est nécessaire d'en préciser les contours épistémologiques :
• Étymologie et intention : Littéralement, la bienveillance consiste à « vouloir du bien à autrui ».
C'est une disposition favorable qui vise la réussite et la réalisation personnelle de l'autre.
• Temporalité (Court terme vs Long terme) : La bienveillance peut impliquer de sacrifier le confort immédiat pour le bien de l'élève à long terme.
Ainsi, la fermeté, l'exigence ou même une sanction peuvent être des actes bienveillants s'ils sont explicités et pratiqués dans le respect de l'élève.
• Distinction fondamentale : Elle ne doit pas être confondue avec :
• Cadre institutionnel : La notion est devenue une valeur centrale de l'Éducation nationale depuis la circulaire de 2014, bien qu'elle fût déjà présente dans le secteur privé et les services publics.
La DGESCO (2013) l'associe à un ensemble d'attitudes physiques, morales et psycho-affectives positives et constantes (respect, confiance, encouragement).
--------------------------------------------------------------------------------
Malgré un consensus apparent, le terme suscite des tensions sur le terrain :
| Type de résistance | Argumentation observée | | --- | --- | | Sentiment de jugement | Certains enseignants perçoivent l'injonction à la bienveillance comme une critique de leurs pratiques passées, sous-entendant qu'ils ne l'auraient pas été auparavant. | | Opposition à l'exigence | Une crainte que l'attention portée au bien-être des élèves ne se fasse au détriment de l'effort nécessaire à la réussite académique. | | Crise de l'autorité | La bienveillance est parfois vue comme une entrave à l'autorité face à des manquements disciplinaires chroniques. | | Complexité systémique | La multiplication des élèves à besoins éducatifs particuliers (EBEP) met les équipes sous pression, rendant la posture bienveillante difficile à maintenir sans formation adéquate. |
--------------------------------------------------------------------------------
La bienveillance se traduit par des actes concrets et une posture éthique dans la relation pédagogique :
La puissance du Feedback
Le levier le plus efficace pour la réussite des élèves est le feedback positif. Il doit être :
• Centré sur l'activité et la méthodologie de l'élève.
• Formulé chaleureusement.
• Porteur de confiance et d'espoir dans les capacités de l'élève.
Le professionnel bienveillant doit être attentif aux signes de fragilité qui peuvent mener au décrochage :
• Signes de découragement ou discours négatif sur l'école.
• Absentéisme et arythmies.
• Sentiments d'insécurité (peur de prendre la parole, honte).
• Mutisme, isolement ou passages fréquents à l'infirmerie/vie scolaire.
Une communication renouvelée
L'horizontalité et l'authenticité sont cruciales pour les nouvelles générations :
• Passer d'un rôle purement académique à une relation de personne à personne.
• Pratiquer l'écoute active (savoir se taire pour laisser l'élève s'exprimer).
• Faire preuve de transparence et de prévisibilité.
--------------------------------------------------------------------------------
L'évaluation est une source majeure de stress (environ 60 % des élèves se disent angoissés par les évaluations). Une évaluation bienveillante implique :
• La suppression de l'implicite.
• Le droit à l'erreur et à la remédiation (possibilité de recommencer).
• Un cadre rassurant qui ne sacrifie pas l'exigence intellectuelle.
La lutte contre le harcèlement (Programme PHARE)
La bienveillance s'incarne dans la création d'une « communauté protectrice » :
• Utilisation de la méthode de la « préoccupation partagée ».
• Recherche d'alternatives à la sanction punitive immédiate pour l'intimidateur, en visant le développement de compétences psychosociales.
La bienveillance passe par une réflexion sur le cadre de vie :
• Création de « jardins zen » ou de salles de calme.
• Réinvention des salles d'étude (espaces de coworking, possibilité de travailler debout ou dans des fauteuils).
• Mise en place de dispositifs permettant le mouvement (ballons, vélos-bureaux) pour favoriser la concentration, notamment des élèves hyperactifs.
--------------------------------------------------------------------------------
La bienveillance ne doit pas être une initiative isolée mais une stratégie d'établissement.
• Le rôle du diagnostic : Utiliser l'auto-évaluation (domaine du climat scolaire et du bien-être) pour identifier les besoins réels des élèves et des familles, en évitant les solutions préconçues.
• La Qualité de Vie au Travail (QVT) : Il existe un lien direct entre le bien-être des personnels et celui des élèves. Un encadrement bienveillant (feedback positif du chef d'établissement, convivialité, confiance déléguée) favorise l'engagement des équipes.
• L'éthique de la rencontre : S'intéresser à la singularité de ce que vit l'élève, au-delà de ses difficultés scolaires. Comme le souligne la sociologie, l'éducation par la rencontre est un levier de raccrochage puissant.
• Le partage et la convivialité : Des actions simples, comme le partage de nourriture (fruits en libre-service, repas de Noël partagé entre agents, élèves et chefs étoilés), peuvent transformer radicalement la relation sociale au sein d'un établissement.
L'autorité et la bienveillance sont compatibles. L'autorité s'exerce de manière éthique lorsqu'elle respecte l'intégrité morale de l'élève.
La sincérité de l'adulte, y compris dans l'expression de ses propres limites ou l'admission d'une erreur, renforce paradoxalement sa légitimité auprès des jeunes.
a system that was potentially self-hosting
self-hosting
eLife Assessment
This study reports an important and novel finding that TENT5A, an enzyme involved in fine-tuning poly(A) tail length on selected mRNAs, is required for proper enamel mineralization in mice. The evidence supporting the authors' conclusion that reduced expression of enamel matrix proteins (EMPs) in TENT5A-deficient mice results from shortened poly(A) tails remains incomplete, as TENT5A may possess additional functions independent of post-transcriptional regulation that are not addressed in the current study.
Reviewer #1 (Public review):
Summary:
The authors aim to determine whether TENT5A, a post-transcriptional regulator previously implicated in bone formation, also plays a role in enamel development. Using a mouse model lacking TENT5A, they report hypomineralized enamel with structural defects, accompanied by reduced expression, altered poly(A) tail length, and impaired secretion of enamel matrix proteins, particularly amelogenin. By combining ultrastructural imaging, transcriptomics, direct RNA sequencing, and protein localization analyses, the study proposes that TENT5A promotes cytoplasmic polyadenylation and translation of a subset of extracellular matrix transcripts required for enamel biomineralization.
Strengths:
A major strength of this work is its conceptual novelty. To my knowledge, this is the first study to demonstrate that a non-canonical poly(A) polymerase plays a direct role in enamel development, extending post-transcriptional regulation by cytoplasmic polyadenylation from bone to enamel, a biologically distinct and non-regenerative mineralized tissue. The identification of amelogenin as a dominant, tissue-specific target provides a new perspective on how enamel matrix production is regulated beyond transcriptional control.
In addition, the study is supported by a comprehensive and complementary set of approaches linking molecular changes to tissue-level phenotypes. The use of direct RNA sequencing provides strong evidence for selective regulation of poly(A) tail length in specific transcripts rather than global effects on mRNA metabolism, and the phenotypic analyses convincingly connect altered post-transcriptional regulation to defects in enamel structure and mineralization.
Weaknesses:
Although the data support a role for TENT5A in stabilizing and promoting translation of amelogenin and related transcripts, the mechanism underlying substrate specificity remains unresolved. Poly(A) tail length alone does not explain why certain transcripts are regulated while others are not, and the proposed involvement of protein partners or RNA processing steps remains speculative. This limitation should be more clearly framed as an open question rather than an emerging mechanism.
A further limitation is the lack of direct human genetic or clinical evidence linking TENT5A to enamel defects. In humans, loss-of-function variants in TENT5A are known to cause a recessive form of osteogenesis imperfecta, but TENT5A has not been associated with amelogenesis imperfecta or other enamel phenotypes. This limits immediate translational interpretation of the mouse enamel phenotype and highlights the need for future human genetic or clinical studies.
Finally, the manuscript does not address whether other members of the TENT5 family are expressed in ameloblasts or could compensate for the loss of TENT5A, leaving open questions about redundancy and specificity within this family.
Reviewer #2 (Public review):
Summary:
The manuscript by Aranaz-Novaliches describes a study of Tent5a knockout (KO) mice. The authors demonstrate a severe enamel phenotype in these mice, characterized by hypoplastic enamel with markedly disturbed organization of enamel rods. Additionally, they report that Amelx expression is reduced in the mutant compared to wild type (WT) at both mRNA and protein levels. The authors also examine the distribution and co-localization of Amelx and Ambn in ameloblasts and the enamel matrix. These findings are novel and provide important insights into the role of polyadenylation in regulating enamel matrix protein translation and its downstream effects on protein trafficking, secretion, and enamel formation. However, I have multiple concerns regarding the data and its analysis that need to be addressed.
Specific comments:
(1) Introduction
The structure of the introduction is unconventional. The first sentence of the third paragraph states that the goal of this study is to investigate the role of TENT5A in enamel formation, but the rest of the paragraph focuses on enamel in general. The following paragraph claims that the authors discovered the effects of Tent5a deficiency on enamel formation for the first time, yet most of the paragraph discusses enamel proteins and amelogenesis. The choice of references is problematic. The authors cite Sire et al. (2007), which focuses on the origin and evolution of enamel mineralisation genes, a poor fit for this context. A more appropriate source would be a recent review, e.g., Lacruz R et al., Physiol Rev. 2017;97(3):939-993. Ambn constitutes ~5% of the enamel matrix, not 10%. Reference 16 (Martin) is not ideal for murine enamel; more detailed studies exist, e.g., Smith CE et al., J Anat. 2019;234(2):274-290. References on protein-protein interactions (17-19) are also off: Wald et al. studied Ambn-Ambn and Amelx-Amelx interactions separately; Fang et al. focused on Amelx self-assembly only; Kawasaki and Weiss addressed gene evolution. The authors should cite work from Moradian-Oldak's lab, which clearly demonstrates Amelx-Ambn interactions. The last paragraph contains confusing statements, e.g., "TENT5a localized in rER promotes the expression of AmelX and other secreted protein transcripts." Also, the manuscript does not convincingly show disruption of self-assembly beyond overall enamel disorganization.
(2) Results
(a) microCT
Quantitative microCT analyses of WT and KO enamel are needed. At a minimum, enamel thickness and density should be measured from at least three biological replicates per genotype. Severe malocclusion in KO mice is not discussed. The mandibular incisor appears abraded, while the maxillary incisor is overgrown. Is maxillary enamel as affected as mandibular? The age of the mice is not specified. High-resolution scans of isolated mandibular incisors described in Materials and Methods should be included.
(b) SEM
The term "disorganized crystal structure" is incorrect - SEM cannot reveal crystal structure. This requires electron/X-ray diffraction or vibrational spectroscopy. Likely, the authors meant disorganized rods and interrod enamel. The phrase "weak HAP composition" is unclear. Can the increase in interprismatic matrix volume and reduction in rod diameter be quantified? Since rods are secreted by distal Tomes' processes and interrod by proximal Tomes' processes, an imbalance may indicate alterations in the ameloblast secretory apparatus. TEM studies of demineralized incisors are recommended to assess ameloblast ultrastructure.
(c) EMP expression
There is a discrepancy between WB images and data in Figure S2a. In Figure 2b, Amelx band is stronger than Ambn (expected, as Amelx is ~20× more abundant), but in Figure S2a, Ambn appears higher. How was protein intensity in Fig. S2a calculated? Optical density? Was normalization applied? Co-localization in Figure 2d was performed on LS8 cells, which lack a true ameloblast phenotype. Amelx expression in LS8 cells is ~2% of actin (Sarkar et al., 2014), whereas in murine incisors, it is ~600× higher than actin (Bui et al., 2023). Ambn signal is weaker than Amelx, which may affect co-localization results.
(d) Splicing products in Figure 2e
All isoforms except one contain exon 4. The major functional splice product of Amelx lacks exon 4 (Haruyama et al. J Oral Biosci. 2011;53(3):257-266), and there are some indications that the presence of exon 4 can lead to enamel defects. Can it be that the observed phenotype is due to the presence of exon 4?
(e) Co-localization studies
The presented co-localization studies do not demonstrate self-assembly defects; they reflect enamel microstructural defects observed by SEM. Self-assembly occurs at the nanoscale and cannot be assessed by light microscopy except with advanced optical methods. Conclusions based on single images are weak. The authors should perform experiments at least on three biological replicates per genotype, quantify results (e.g., total gray values per ROI of equal pixel size), and use co-localization metrics such as Mander's coefficient. Claims about alternative secretory pathways require much stronger evidence.
The authors should avoid implying that mRNA is inside the ER lumen. It is likely associated with the outer rER surface, which is expected. The resolution of the methods used is insufficient to confirm ER lumen localization.
Reviewer #3 (Public review):
Summary:
It is well established that poly(A) tails at the 3' end of mRNA are critical for mRNA stability, providing another layer of gene regulation. TENT5A is one of the non-canonical poly(A) polymerases that add an extra poly(A) tail. This manuscript demonstrates that the Tent5A mutation leads to mineralization abnormalities in the tooth, shorter poly(A) tails in amelogenin mRNA and some other selected mRNAs, and provides a list of TENT5A interacting proteins.
Strengths:
(1) The authors show in vivo genetic evidence that Tent5a is critical for normal tooth mineralization.
(2) The authors show that the length of the poly(A) tail in amelogenin (AmelX) is 13 bases shorter in Tent5a mutants but not in other mRNAs, such as ameloblastin (Ambn).
(3) Differentially expressed genes (DEGs) in Tent5A mutant tissues (cervical loop) are identified, and some of them show different lengths of poly(A) tails.
(4) TENT5A interacting proteins are identified. Together with the DEGs, these datasets will provide valuable research tools to the community.
Weaknesses:
(1) There is no direct evidence to support the main conclusion; the length of the poly(A) tail is critical for normal tooth mineralization.
(2) The RNAseq data to identify TENT5A substrate is based on the assumption that shorter poly(A) tailed RNA is less stable. However, there are multiple reasons for the differential expression of RNA in Tent5A mutant tissues.
(3) Several TENT5A-interacting proteins have been identified, but, beyond their colocalization with a target mRNA, no mechanistic studies have been conducted.
Author response:
We thank the editors and reviewers for their careful and constructive evaluation of our manuscript. We appreciate the recognition of the conceptual novelty and in vivo relevance of our findings. We have carefully considered all comments and outline below the major revisions and additional analyses we will undertake. For clarity, we address the reviewers’ comments in thematic sections.
Cell-autonomous contribution of Tent5a to phenotype
We agree that the use of a complete knockout model raises the possibility of indirect or non-cell-autonomous effects on tooth development, particularly given the observed dentin alterations. To address this point directly, we are generating and analyzing an ameloblast-specific conditional model we have already on shelf (Ambn-Cre; Tent5a<sup>flox/flox</sup>) to determine whether the enamel phenotype arises from cell-autonomous loss of TENT5A in the secretory epithelium. This approach will allow us to distinguish epithelial-intrinsic effects from potential secondary contributions of odontoblasts or mesenchymal tissues. Results from this model will be incorporated into the revised manuscript.
Mechanistic basis and substrate specificity
We agree that the mechanism underlying substrate selectivity of TENT5A requires further clarification. We have performed multiple classical RNA–protein interaction assays, including CLIP-based approaches, without identifying a clear sequence-specific recognition motif. In the revised manuscript, we will present substrate specificity as an open mechanistic question rather than implying a defined recognition mechanism.
To strengthen this aspect, we will extend our analysis to include combined immunoprecipitation strategies and investigation of potential ribosome-associated or co-translational interactions of TENT5A.
In addition, we will further validate selected high-confidence TENT5A interactors identified in our dataset in context of putative changes in AmelX-polyA tail length.
Poly(A) tail length and functional causality
We acknowledge that shortening of the poly(A) tail alone does not formally establish causality. However, our data consistently show that TENT5A-dependent shortening of poly(A) tails correlates with reduced mRNA and protein levels of key enamel matrix components. In the revised manuscript, we will clarify this mechanistic framework more explicitly, integrating poly(A) length, transcript abundance, and protein-level data in a structured manner, while clearly distinguishing correlation from formal proof of causality.
We will also perform additional functional assays, including mRNA stability measurements in vitro in cells with genetic ablation of Tent5a, to further test the link between poly(A) shortening and reduced AmelX protein levels.
Quantitative microCT and enamel morphology
We will include quantitative microCT analyses of enamel thickness and mineral density from multiple biological replicates per genotype (n ≥ 3). Sample numbers will be explicitly stated throughout. Additional high-resolution scans of isolated incisors will be provided. We will also quantify occlusal angle and include whole-skull reconstructions to document malocclusion. Maxillary enamel will be analyzed and quantified alongside mandibular enamel.
SEM terminology will be corrected (e.g., replacing “crystal structure” with “rod/interrod organization”), and structural parameters such as rod diameter and interprismatic matrix proportion will be quantitatively assessed.
We agree that ultrastructural analysis of ameloblast secretory morphology is important. We have experience with TEM analysis of demineralized incisors and will perform additional ultrastructural examination to assess the integrity of Tomes’ processes and the secretory apparatus in Tent5a-deficient ameloblasts. These data will allow us to distinguish between primary alterations in secretory morphology and downstream effects on matrix organization.
Amelx splice variants
We will re-analyze our RNA-seq data with specific attention to exon 4-containing isoforms and clarify the distribution of splice variants in WT and KO samples. These findings will be explicitly discussed in the context of prior literature.
Co-localization and self-assembly claims
We agree that conventional light microscopy cannot directly resolve nanoscale self-assembly events. In Figure 3, our intention was to demonstrate differential subcellular distribution and partial segregation of AMELX and AMBN within secretory compartments, rather than to claim direct visualization of molecular self-assembly. In the revised manuscript, we will clarify this distinction, moderate the terminology accordingly, and provide explicit quantitative co-localization analyses across multiple biological replicates.
TENT5 family paralogs
To address potential redundancy within the TENT5 family, we will analyze published single-cell RNA-seq datasets (Sharir et al., 2019; Krivanek et al., 2020) to assess expression of TENT5 paralogs in ameloblasts. These findings will be validated using targeted transcriptional analyses.
Human clinical relevance
We appreciate the suggestion to examine potential human enamel phenotypes. We will pursue retrospective analysis of clinical and imaging data from patients carrying TENT5A variants through our collaborations with rare disease networks and specialized centers in Europe and the United States. Any relevant findings will be incorporated into the revised manuscript.
Tissue sampling clarification
We apologize for imprecise terminology regarding transcriptomic sampling. The analyzed tissue corresponds to the proximal incisor region up to the mineralization stage. We will include a schematic and clarify nomenclature throughout the manuscript.
Language and data clarity
The manuscript will be thoroughly revised for clarity, consistency of terminology, figure referencing, and accuracy of citations. We will explicitly clarify the methodology used for protein quantification, including normalization strategy and densitometric analysis, to address inconsistencies noted in the supplementary data. We will also expand the discussion to address the biological relevance of moderate poly(A) shortening, referencing established literature demonstrating that even subtle changes in tail length can significantly influence translational efficiency.
Although AMELX is the most abundant enamel matrix protein and exhibits a consistent TENT5A-dependent poly(A) shortening phenotype, our data demonstrate that multiple secreted proteins are similarly affected. We will revise the text to clearly articulate that the enamel phenotype likely reflects the combined contribution of multiple TENT5A-regulated secretory factors rather than a single-gene effect.
We believe these revisions will substantially strengthen the mechanistic, quantitative, and conceptual framework of the study and provide a clearer foundation for interpreting TENT5A-dependent regulation of enamel biomineralization.
Note de Synthèse : Les Enjeux de la Professionnalité Enseignante et de l'Éthique Relationnelle
Ce document synthétise les interventions du webinaire du 12 avril 2023, animé par Christophe Marsollier, Inspecteur général de l'éducation, du sport et de la recherche.
L'analyse explore la mutation profonde du métier d'enseignant face à la complexité croissante du milieu scolaire. Les points de bascule majeurs identifiés incluent l'intégration systémique des compétences psychosociales (CPS), l'adoption de pédagogies institutionnelles et coopératives, et le passage d'une posture de « sachant » à celle d'« écoutant ».
La réussite de l'élève est ici pensée non seulement par la transmission académique, mais par une « éthique relationnelle » fondée sur la confiance, la reconnaissance de la vulnérabilité et le bien-être eudémonique.
--------------------------------------------------------------------------------
Le développement des CPS n'est plus considéré comme une activité périphérique, mais comme la « trame de fond » de la réussite scolaire et de la santé mentale.
Les CPS englobent les capacités permettant à l’élève de :
• Gérer ses émotions : Appréhender son ressenti et celui d'autrui.
• S'auto-réguler : Gérer les conflits en autonomie et appartenir au groupe.
• Développer sa citoyenneté : Favoriser le dialogue, l’échange et la collaboration.
Les témoignages d'enseignants (notamment du réseau Jean Lolive à Pantin) soulignent une transformation concrète :
• Apaisement du climat : Moins de besoin de « faire le gendarme » ; les élèves règlent les conflits en amont.
• Disponibilité cognitive : Des élèves sereins et empathiques sont plus aptes à entrer dans les apprentissages pédagogiques.
• Transformation de l'enseignant : Le passage d'un scepticisme initial à une confiance réelle en la capacité d'agir des élèves.
Une instruction interministérielle, rédigée par Santé Publique France, fixe un horizon à 2037 pour la formation généralisée de la population aux CPS.
• Cadre de référence : Publication d'un référentiel en février 2022 pour uniformiser les pratiques.
• Pédagogies préconisées : Utilisation de méthodes expérientielles comme le « théâtre forum » ou le « jeu des trois figures » pour favoriser le changement de rôle et l'empathie.
--------------------------------------------------------------------------------
L'analyse met en lumière la pédagogie institutionnelle (PI) comme levier de transformation, particulièrement en éducation prioritaire (REP+).
Inspiré du mouvement Freinet, ce dispositif permet d'articuler éducation (épanouissement) et transmission (connaissances).
| Caractéristique | Fonctionnement et Bénéfices | | --- | --- | | Différenciation | L'élève choisit son niveau de ceinture par compétence (ex: se repérer dans le temps). | | Auto-évaluation | Utilisation de fichiers autocorrectifs ; l'élève identifie ses stratégies d'apprentissage. | | Évaluation Positive | La ceinture sanctionne la réussite (100% requis) et non le manque. L'échec est une étape formative. | | Coopération | Les élèves ayant validé des ceintures deviennent des « aides » pour leurs pairs. |
La classe est pensée comme une « petite société » régulée par :
• Le Conseil : Lieu de décision collective où l'on critique l'organisation et propose des améliorations.
• Les Responsabilités : Métiers spécifiques (responsable du temps, du matériel, des affichages) qui donnent une place à chacun.
• L'Espace de parole : Le « Quoi de neuf » et les temps de météo intérieure pour intégrer les affects.
--------------------------------------------------------------------------------
Le métier d'enseignant est comparé à celui d'un tisseur, créateur de liens et de sens dans un monde « déchiré ».
S'appuyant sur les travaux d'Edgar Morin et d'Adrien Rivard, Christophe Marsollier évoque la nécessité de cultiver :
Pour maintenir le « tissu » de la relation pédagogique, quatre piliers sont identifiés :
• La Confiance : Elle doit être mutuelle et engagée.
• Le Respect : L'élève doit être considéré comme une personne à part entière, avec sa dignité propre.
• L'Écoute : Sortir de l'écoute passive pour une écoute active des difficultés et erreurs.
• Le Droit à l'erreur : Admis pour l'élève, mais aussi pour l'enseignant qui doit pouvoir s'excuser.
--------------------------------------------------------------------------------
L'enseignement, particulièrement en milieu défavorisé, exige une attention particulière à la vulnérabilité des élèves.
• La Blessure Psychologique : Les élèves en grande vulnérabilité sont souvent touchés dans leurs besoins fondamentaux (sécurité, reconnaissance, justice).
• L'Intérêt pour la Singularité : Les recherches (Virginie Muniglia) montrent que le besoin premier des jeunes vulnérables est que l'adulte s'intéresse à leur singularité, et non qu'il les traite de manière standardisée.
• Le Tact Pédagogique : Capacité (théorisée par Éric Prairat) à trouver le bon geste et le bon mot au bon moment, en s'adaptant à l'imprévu.
--------------------------------------------------------------------------------
Le document clarifie la notion de bien-être, souvent mal comprise dans le cadre scolaire.
• Bien-être Hédonique : Recherche du plaisir immédiat (souvent critiqué à l'école).
• Bien-être Eudémonique : Sentiment de satisfaction ressenti lorsqu'on est captivé par une activité, qu'on dépasse une difficulté ou que l'on progresse.
C'est le « bien-être optimal » ou l'expérience autotélique.
Le bien-être n'est pas le but ultime de l'école, mais la condition indispensable pour favoriser la réussite, notamment pour les élèves les plus fragiles.
Il permet de transformer la « souffrance relationnelle » en un climat d'exigence bienveillante.
--------------------------------------------------------------------------------
En conclusion, le métier d'enseignant est décrit comme une profession de la relation, nécessitant :
• Le Travail Collectif : Créer des communautés d'apprentissage professionnel pour rompre l'isolement et penser la pratique (monographies, analyses de pratiques).
• La Foi en l'Éducabilité : Posture philosophique (Philippe Meirieu) consistant à croire inconditionnellement en la capacité de chaque élève à progresser.
• La Joie comme Boussole : Pour les jeunes enseignants, le critère de la joie et de l'alignement personnel est présenté comme le meilleur garant de la créativité et de l'efficacité pédagogique.
« La question n'est pas quel monde laisserons-nous à nos enfants, mais quels enfants laisserons-nous au monde. » — Philippe Meirieu (cité en conclusion).
RRID:CVCL_1258
DOI: 10.1158/0008-5472.CAN-25-2086
Resource: (KCB Cat# KCB 2014032YJ, RRID:CVCL_1258)
Curator: @evieth
SciCrunch record: RRID:CVCL_1258
RRID:CVCL_0553
DOI: 10.1158/0008-5472.CAN-25-2086
Resource: (RRID:CVCL_0553)
Curator: @evieth
SciCrunch record: RRID:CVCL_0553
Addgene # 133861
DOI: 10.1111/febs.70278
Resource: RRID:Addgene_133861
Curator: @evieth
SciCrunch record: RRID:Addgene_133861
RRID:CVCL_0030
DOI: 10.1111/febs.70278
Resource: (ICLC Cat# HTL95023, RRID:CVCL_0030)
Curator: @evieth
SciCrunch record: RRID:CVCL_0030
Addgene #112865
DOI: 10.1038/s44321-025-00369-2
Resource: RRID:Addgene_112865
Curator: @evieth
SciCrunch record: RRID:Addgene_112865
CRL-3216
DOI: 10.1038/s44321-025-00369-2
Resource: (RRID:CVCL_0063)
Curator: @evieth
SciCrunch record: RRID:CVCL_0063
RRID:SCR_023747
DOI: 10.1038/s44321-025-00364-7
Resource: Zeiss Zen Lite (RRID:SCR_023747)
Curator: @evieth
SciCrunch record: RRID:SCR_023747
RRID:SCR_002285
DOI: 10.1038/s44321-025-00352-x
Resource: Fiji (RRID:SCR_002285)
Curator: @evieth
SciCrunch record: RRID:SCR_002285
RRID:SCR_023543
DOI: 10.1038/s44321-025-00352-x
Resource: Harmony (RRID:SCR_023543)
Curator: @evieth
SciCrunch record: RRID:SCR_023543
Addgene #85140
DOI: 10.1038/s44321-025-00352-x
Resource: RRID:Addgene_85140
Curator: @evieth
SciCrunch record: RRID:Addgene_85140
Addgene #12260
DOI: 10.1038/s44321-025-00352-x
Resource: RRID:Addgene_12260
Curator: @evieth
SciCrunch record: RRID:Addgene_12260
RRID: CVCL_2544
DOI: 10.1038/s42003-026-09518-w
Resource: (CLS Cat# 300433/p2464_KHOS-240S, RRID:CVCL_2544)
Curator: @evieth
SciCrunch record: RRID:CVCL_2544
RRID: CVCL_0439
DOI: 10.1038/s42003-026-09518-w
Resource: (KCLB Cat# 21543, RRID:CVCL_0439)
Curator: @evieth
SciCrunch record: RRID:CVCL_0439
ATCC CCL-81
DOI: 10.1016/j.xpro.2026.104367
Resource: (IZSLER Cat# BS CL 86, RRID:CVCL_0059)
Curator: @evieth
SciCrunch record: RRID:CVCL_0059
ATCC CRL-3276
DOI: 10.1016/j.xpro.2026.104367
Resource: (ATCC Cat# CRL-3276, RRID:CVCL_DA06)
Curator: @evieth
SciCrunch record: RRID:CVCL_DA06
ATCC CRL-4000
DOI: 10.1016/j.xpro.2026.104367
Resource: (ATCC Cat# CRL-4000, RRID:CVCL_4388)
Curator: @evieth
SciCrunch record: RRID:CVCL_4388
Jackson LabsCat# 008169
DOI: 10.1016/j.xcrm.2026.102631
Resource: (IMSR Cat# JAX_008169,RRID:IMSR_JAX:008169)
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:008169
Jackson LabsCat# 000664
DOI: 10.1016/j.xcrm.2026.102631
Resource: RRID:IMSR_JAX:000664
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:000664
RRID:CVCL_0045
DOI: 10.1016/j.xcrm.2026.102631
Resource: (DSMZ Cat# ACC-305, RRID:CVCL_0045)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0045
ATCCCRL-3216
DOI: 10.1016/j.str.2026.01.011
Resource: (RRID:CVCL_0063)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0063
RRID:CVCL_6926
DOI: 10.1016/j.redox.2026.104073
Resource: (RRID:CVCL_6926)
Curator: @evieth
SciCrunch record: RRID:CVCL_6926
ATCCCRL-11268
DOI: 10.1016/j.neuron.2025.12.028
Resource: (RRID:CVCL_1926)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1926
RRID: IMSR_JAX: 003010
DOI: 10.1016/j.neuron.2025.12.021
Resource: (IMSR Cat# JAX_003010,RRID:IMSR_JAX:003010)
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:003010
RRID: CVCL_1045
DOI: 10.1016/j.neo.2026.101289
Resource: (ECACC Cat# 05092802, RRID:CVCL_1045)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1045
ATCCCat#HTB-38
DOI: 10.1016/j.molcel.2026.01.018
Resource: (RRID:CVCL_0320)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0320
ATCCCCL-247
DOI: 10.1016/j.molcel.2026.01.012
Resource: (RRID:CVCL_VU38)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_VU38
ATCCCCL-121
DOI: 10.1016/j.molcel.2026.01.012
Resource: (BCRJ Cat# 0110, RRID:CVCL_0317)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0317
ATCCCRL-3216
DOI: 10.1016/j.molcel.2026.01.012
Resource: (RRID:CVCL_0063)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0063
ATCCCCL-228
DOI: 10.1016/j.molcel.2026.01.012
Resource: (KCB Cat# KCB 200848YJ, RRID:CVCL_0546)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0546
ATCCCCL-185
DOI: 10.1016/j.molcel.2026.01.012
Resource: (CCLV Cat# CCLV-RIE 1035, RRID:CVCL_0023)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0023
ATCCCRL-6322
DOI: 10.1016/j.molcel.2026.01.012
Resource: (BCRJ Cat# 0044, RRID:CVCL_0604)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0604
AddgenePlasmid #12260
DOI: 10.1016/j.molcel.2026.01.012
Resource: RRID:Addgene_12260
Curator: @areedewitt04
SciCrunch record: RRID:Addgene_12260
ATCCCCL-243
DOI: 10.1016/j.molcel.2026.01.012
Resource: (KCB Cat# KCB 90029YJ, RRID:CVCL_0004)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0004
ATCCTIB-152
DOI: 10.1016/j.molcel.2026.01.012
Resource: (NIH-ARP Cat# 177-450, RRID:CVCL_0367)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0367
ATCCHTB-96
DOI: 10.1016/j.molcel.2026.01.012
Resource: (RRID:CVCL_0042)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0042
ATCCCCL-2
DOI: 10.1016/j.molcel.2026.01.012
Resource: (TKG Cat# TKG 0331, RRID:CVCL_0030)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0030
ATCCCCL-221
DOI: 10.1016/j.molcel.2026.01.012
Resource: (RRID:CVCL_0248)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0248
Python
DOI: 10.1016/j.isci.2026.115085
Resource: Python Programming Language (RRID:SCR_008394)
Curator: @areedewitt04
SciCrunch record: RRID:SCR_008394
Python
DOI: 10.1016/j.isci.2026.115012
Resource: Python Programming Language (RRID:SCR_008394)
Curator: @areedewitt04
SciCrunch record: RRID:SCR_008394
Charles River Strain code: 027
DOI: 10.1016/j.isci.2026.115008
Resource: RRID:IMSR_CRL:027
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_CRL:027
CRL-1658
DOI: 10.1016/j.isci.2026.114825
Resource: (RCB Cat# RCB2767, RRID:CVCL_0594)
Curator: @evieth
SciCrunch record: RRID:CVCL_0594
CRL-1446
DOI: 10.1016/j.isci.2026.114825
Resource: (ATCC Cat# CRL-1446, RRID:CVCL_0286)
Curator: @evieth
SciCrunch record: RRID:CVCL_0286
RRID: CVCL_0511
DOI: 10.1016/j.intimp.2026.116295
Resource: (ATCC Cat# TIB-71, RRID:CVCL_0493)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0493
ATCCCat# CCL-2
DOI: 10.1016/j.immuni.2025.12.017
Resource: (ICLC Cat# HTL95023, RRID:CVCL_0030)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0030
The Jackson LaboratoryJAX-000664
DOI: 10.1016/j.devcel.2026.01.015
Resource: RRID:IMSR_JAX:000664
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:000664
ATCCCat# CRL-1682
DOI: 10.1016/j.crmeth.2026.101307
Resource: (NCBI_Iran Cat# C558, RRID:CVCL_0152)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0152
ATCCCat# CRL-1469
DOI: 10.1016/j.crmeth.2026.101307
Resource: (ECACC Cat# 87092802, RRID:CVCL_0480)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0480
ATCCCat# HTB-26
DOI: 10.1016/j.crmeth.2026.101307
Resource: (RRID:CVCL_0062)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0062
ATCCCat# HTB-161
DOI: 10.1016/j.crmeth.2026.101307
Resource: (ATCC Cat# HTB-161, RRID:CVCL_0465)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0465
Jackson Laboratory#000664
DOI: 10.1016/j.cmet.2026.01.019
Resource: (IMSR Cat# JAX_000664,RRID:IMSR_JAX:000664)
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:000664
ATCCCRL-6475
DOI: 10.1016/j.cmet.2026.01.015
Resource: (KCLB Cat# 80008, RRID:CVCL_0159)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0159
ATCCCRL-11268
DOI: 10.1016/j.cmet.2026.01.015
Resource: (RRID:CVCL_1926)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1926
RRID: CVCL_0007
DOI: 10.1016/j.celrep.2026.117010
Resource: (JCRB Cat# IFO50038, RRID:CVCL_0007)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0007
RRID: CVCL_2120
DOI: 10.1016/j.celrep.2026.117010
Resource: (DSMZ Cat# ACC-555, RRID:CVCL_2120)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_2120
RRID: CVCL_1609
DOI: 10.1016/j.celrep.2026.117010
Resource: (RRID:CVCL_1609)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1609
Jackson Laboratory#002014
DOI: 10.1016/j.celrep.2026.117010
Resource: (IMSR Cat# JAX_002014,RRID:IMSR_JAX:002014)
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:002014
Jackson Laboratory#000664
DOI: 10.1016/j.celrep.2026.117010
Resource: RRID:IMSR_JAX:000664
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:000664
Addgene # 12371
DOI: 10.1016/j.celrep.2026.117010
Resource: RRID:Addgene_12371
Curator: @areedewitt04
SciCrunch record: RRID:Addgene_12371
KerafastEWL001-FP
DOI: 10.1016/j.celrep.2026.116994
Resource: (RRID:CVCL_ZD32)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_ZD32
Jackson Labsstock #000664
DOI: 10.1016/j.celrep.2026.116994
Resource: RRID:IMSR_JAX:000664
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_JAX:000664
KerafastEWL002-FP
DOI: 10.1016/j.celrep.2026.116994
Resource: (RRID:CVCL_ZD33)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_ZD33
Millipore SigmaSCC193
DOI: 10.1016/j.celrep.2026.116994
Resource: (TKG Cat# TKG 0484, RRID:CVCL_1288)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1288
AddgeneCat#100097
DOI: 10.1016/j.celrep.2026.116992
Resource: RRID:Addgene_100097
Curator: @areedewitt04
SciCrunch record: RRID:Addgene_100097
AddgeneCat#60032
DOI: 10.1016/j.celrep.2026.116992
Resource: RRID:Addgene_60032
Curator: @areedewitt04
SciCrunch record: RRID:Addgene_60032
ZDB-GENO-960809-7
DOI: 10.1016/j.celrep.2026.116971
Resource: RRID:ZFIN_ZDB-GENO-960809-7
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-GENO-960809-7
Jackson LaboratoryJIPSC001000
DOI: 10.1016/j.celrep.2026.116971
Resource: (RRID:CVCL_B5P3)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_B5P3
ATCCACS-1030
DOI: 10.1016/j.celrep.2026.116971
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0A07
ZFIN: ZDB-ALT-050916-14
DOI: 10.1016/j.celrep.2026.116971
Resource: (ZFIN Cat# ZDB-ALT-050916-14,RRID:ZFIN_ZDB-ALT-050916-14)
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-050916-14
ATCCACS-1026
DOI: 10.1016/j.celrep.2026.116971
Resource: (RRID:CVCL_0A05)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0A05
ZFIN: ZDB-ALT-060821-4
DOI: 10.1016/j.celrep.2026.116971
Resource: (ZFIN Cat# ZDB-ALT-060821-4,RRID:ZFIN_ZDB-ALT-060821-4)
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-060821-4
ZFIN: ZDB-ALT-120723-3
DOI: 10.1016/j.celrep.2026.116971
Resource: (ZFIN Cat# ZDB-ALT-120723-3,RRID:ZFIN_ZDB-ALT-120723-3)
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-120723-3
ZFIN: ZDB-ALT-110411-1
DOI: 10.1016/j.celrep.2026.116971
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-110411-1
ZFIN: ZDB-ALT-040601-2
DOI: 10.1016/j.celrep.2026.116971
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-040601-2
ZFIN: ZDB-ALT-120828-2
DOI: 10.1016/j.celrep.2026.116971
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-120828-2
RRID: CVCL_1056
DOI: 10.1016/j.celrep.2026.116968
Resource: (RRID:CVCL_1056)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1056
ATCCCRL-1619
DOI: 10.1016/j.cell.2026.01.009
Resource: (BCRJ Cat# 0278, RRID:CVCL_0132)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0132
ATCCCRL-3216
DOI: 10.1016/j.cell.2026.01.009
Resource: (RRID:CVCL_0063)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0063
ATCCCRL-5803
DOI: 10.1016/j.cell.2026.01.009
Resource: (NCI-DTP Cat# NCI-H1299, RRID:CVCL_0060)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0060
Addgene; Subach et al.49Cat#55294
DOI: 10.1016/j.cell.2026.01.009
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:Addgene_55294
Charles River088
DOI: 10.1016/j.cell.2026.01.009
Resource: (IMSR Cat# CRL_088,RRID:IMSR_CRL:088)
Curator: @areedewitt04
SciCrunch record: RRID:IMSR_CRL:088
ATCCHTB-96
DOI: 10.1016/j.cell.2026.01.009
Resource: (RRID:CVCL_0042)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0042
ATCCCCL-185
DOI: 10.1016/j.cell.2026.01.009
Resource: (CCLV Cat# CCLV-RIE 1035, RRID:CVCL_0023)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0023
ATCC) (CRL-3216
DOI: 10.1016/j.celrep.2026.116990
Resource: (RRID:CVCL_0063)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0063
RRID: CVCL_E2RM
DOI: 10.1016/j.cej.2026.174157
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_E2RM
RRID: CVCL_0493
DOI: 10.1016/j.cej.2026.174137
Resource: (ATCC Cat# TIB-71, RRID:CVCL_0493)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0493
RRID: CVCL_0063
DOI: 10.1016/j.canlet.2026.218328
Resource: (RRID:CVCL_0063)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0063
RRID: CVCL_1092
DOI: 10.1016/j.canlet.2026.218328
Resource: (NCI-DTP Cat# BT-549, RRID:CVCL_1092)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_1092
RRID: CVCL_0062
DOI: 10.1016/j.canlet.2026.218328
Resource: (RRID:CVCL_0062)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0062
RRID: CVCL_0553
DOI: 10.1016/j.canlet.2026.218328
Resource: (RRID:CVCL_0553)
Curator: @areedewitt04
SciCrunch record: RRID:CVCL_0553
RRID:CVCL_0566
DOI: 10.1007/s00109-026-02650-4
Resource: (BCRJ Cat# 0240, RRID:CVCL_0566)
Curator: @evieth
SciCrunch record: RRID:CVCL_0566
RRID:CVCL_2078
DOI: 10.1007/s00109-026-02650-4
Resource: (DSMZ Cat# ACC-541, RRID:CVCL_2078)
Curator: @evieth
SciCrunch record: RRID:CVCL_2078
RRID:CVCL_E332
DOI: 10.1007/s00109-026-02650-4
Resource: (RRID:CVCL_E332)
Curator: @evieth
SciCrunch record: RRID:CVCL_E332
RRID:CVCL_2989
DOI: 10.1007/s00109-026-02650-4
Resource: (JCRB Cat# JCRB1179, RRID:CVCL_2989)
Curator: @evieth
SciCrunch record: RRID:CVCL_2989
RRID:CVCL_J431
DOI: 10.1007/s00109-026-02650-4
Resource: (RRID:CVCL_J431)
Curator: @evieth
SciCrunch record: RRID:CVCL_J431
RRID:CVCL_1357
DOI: 10.1007/s00109-026-02650-4
Resource: (DSMZ Cat# ACC-49, RRID:CVCL_1357)
Curator: @evieth
SciCrunch record: RRID:CVCL_1357
MMRRC_032108‐UCD
DOI: 10.1002/advs.202513670
Resource: (MMRRC Cat# 032108-UCD,RRID:MMRRC_032108-UCD)
Curator: @AleksanderDrozdz
SciCrunch record: RRID:MMRRC_032108-UCD
RRID:SCR_010279
DOI: 10.7554/eLife.108259
Resource: Adobe Illustrator (RRID:SCR_010279)
Curator: @scibot
SciCrunch record: RRID:SCR_010279
RRID:SCR_002285
DOI: 10.7554/eLife.108259
Resource: Fiji (RRID:SCR_002285)
Curator: @scibot
SciCrunch record: RRID:SCR_002285
RRID:SCR_007370
DOI: 10.7554/eLife.108259
Resource: Imaris (RRID:SCR_007370)
Curator: @scibot
SciCrunch record: RRID:SCR_007370
RRID:BDSC_26669
DOI: 10.7554/eLife.108259
Resource: RRID:BDSC_26669
Curator: @scibot
SciCrunch record: RRID:BDSC_26669
RRID:SCR_002798
DOI: 10.7554/eLife.108259
Resource: GraphPad Prism (RRID:SCR_002798)
Curator: @scibot
SciCrunch record: RRID:SCR_002798
RRID:AB_2335286
DOI: 10.7554/eLife.108259
Resource: (Thermo Fisher Scientific Cat# ICN55976, RRID:AB_2335286)
Curator: @scibot
SciCrunch record: RRID:AB_2335286
RRID:BDSC_35785
DOI: 10.7554/eLife.108259
Resource: RRID:BDSC_35785
Curator: @scibot
SciCrunch record: RRID:BDSC_35785