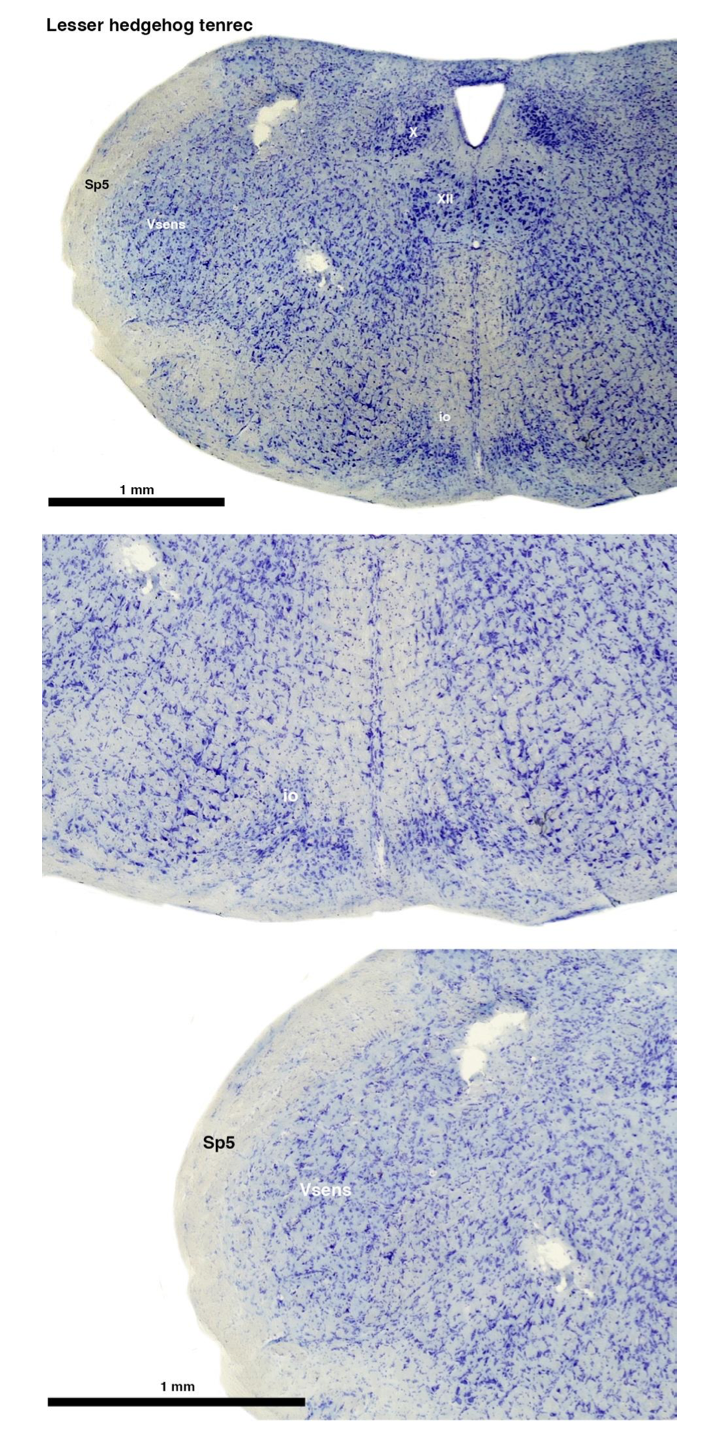
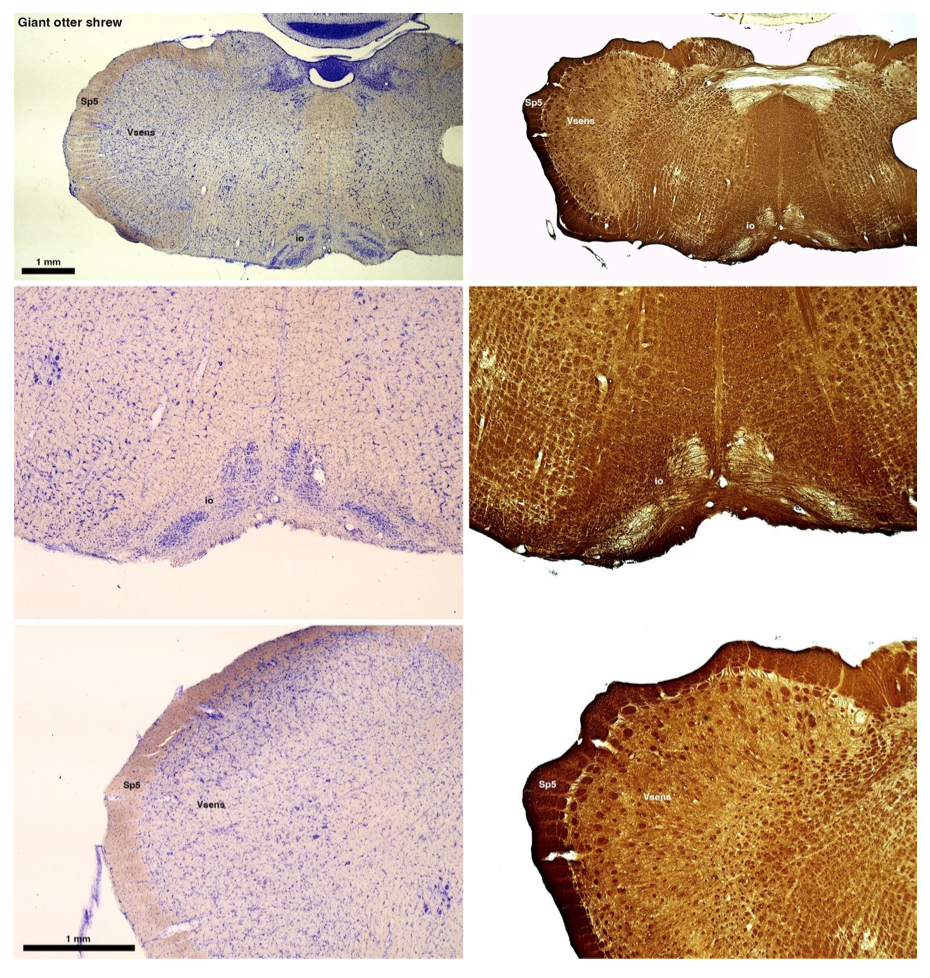
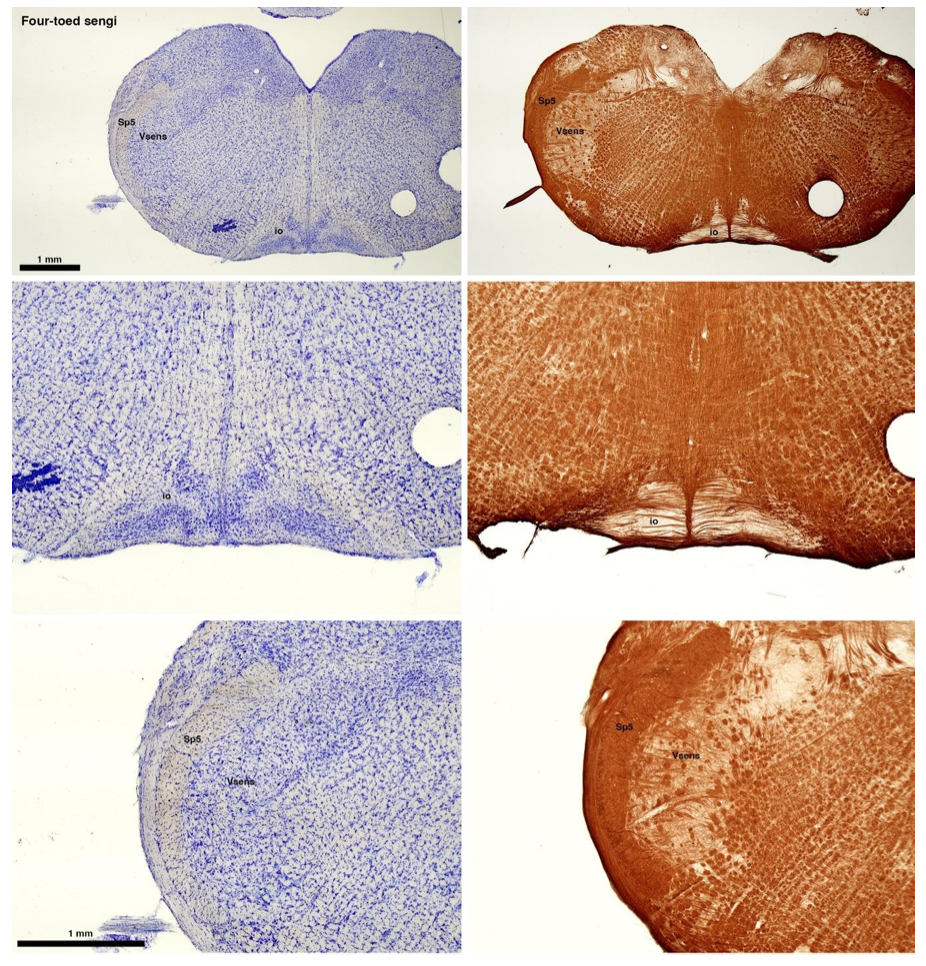
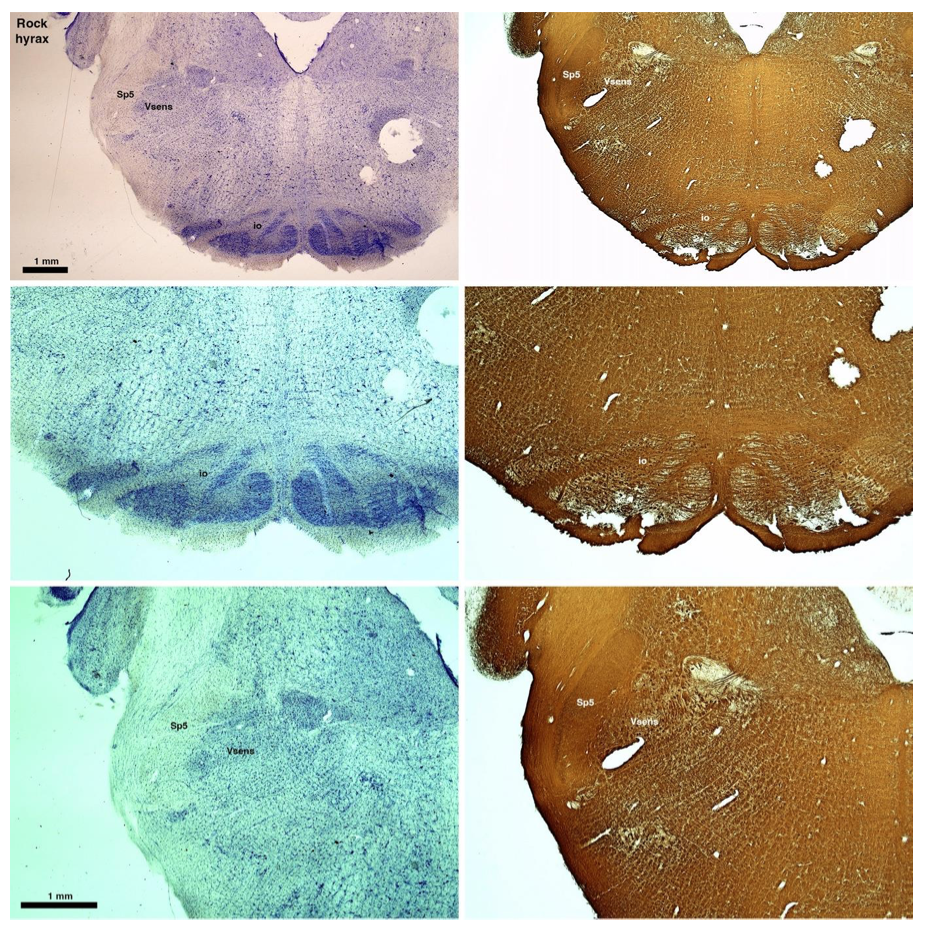
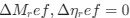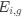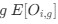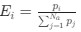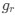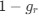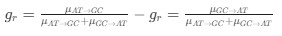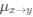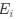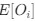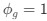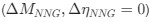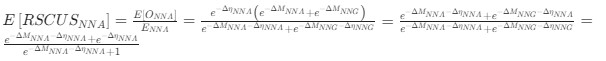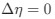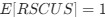Reviewer #2 (Public review):
Summary:
This work by den Bakker and Kloosterman contributes to the vast body of research exploring the dynamics governing the communication between the hippocampus (HPC) and the medial prefrontal cortex (mPFC) during spatial learning and navigation. Previous research showed that population activity of mPFC neurons is replayed during HPC sharp-wave ripple events (SWRs), which may therefore correspond to privileged windows for the transfer of learned navigation information from the HPC, where initial learning occurs, to the mPFC, which is thought to store this information long term. Indeed, it was also previously shown that the activity of mPFC neurons contains task-related information that can inform about the location of an animal in a maze, which can predict the animals' navigational choices. Here, the authors aim to show that the mPFC neurons that are modulated by HPC activity (SWRs and theta rhythms) are distinct from those "encoding" spatial information. This result could suggest that the integration of spatial information originating from the HPC within the mPFC may require the cooperation of separate sets of neurons.
This observation may be useful to further extend our understanding of the dynamics regulating the exchange of information between the HPC and mPFC during learning. However, my understanding is that this finding is mainly based upon a negative result, which cannot be statistically proven by the failure to reject the null hypothesis. Moreover, in my reading, the rest of the paper mainly replicates phenomena that have already been described, with the original reports not correctly cited. My opinion is that the novel elements should be precisely identified and discussed, while the current phrasing in the manuscript, in most cases, leads readers to think that these results are new. Detailed comments are provided below.
Major concerns:
ORIGINAL COMMENT: (1) The main claim of the manuscript is that the neurons involved in predicting upcoming choices are not the neurons modulated by the HPC. This is based upon the evidence provided in Figure 5, which is a negative result that the authors employ to claim that predictive non-local representations in the mPFC are not linked to hippocampal SWRs and theta phase. However, it is important to remember that in a statistical test, the failure to reject the null hypothesis does not prove that the null hypothesis is true. Since this claim is so central in this work, the authors should use appropriate statistics to demonstrate that the null hypothesis is true. This can be accomplished by showing that there is no effect above some size that is so small that it would make the effect meaningless (see https://doi.org/10.1177/070674370304801108).
AUTHOR RESPONSE: We would like to highlight a few important points here. (1) We indeed do not intend to claim that the SWR-modulated neurons are not at all involved in predicting upcoming choice, just that the SWR-unmodulated neurons may play a larger role. We have rephrased the title and abstract to make this clearer.
REVIEWER COMMENT: The title has been rephrased but still conveys the same substantive claim. The abstract sentence also does not clearly state what was found. Using "independently" in the new title continues to imply that SWR modulation and prediction of upcoming choices are separate phenomena. By contrast, in your response here in the rebuttall you state only that "SWR-unmodulated neurons may play a larger role," which is a much more tempered claim than what the manuscript currently argues. Why is this clarification not adopted in the article? Moreover, the main text continues to use the same arguments as before; beyond the cosmetic changes of title and abstract, the claim itself has not materially changed.
AUTHOR RESPONSE: (2) The hypothesis that we put forward is based not only on a negative effect, but on the findings that: the SWR-unmodulated neurons show higher spatial tuning (Fig 3b), more directional selectivity (Fig 3d), more frequent encoding of the upcoming choice at the choice point (new analysis, added in Fig 4d), and higher spike rates during the representations of the upcoming choice (Fig 5b). This is further highlighted by the fact that the representations of upcoming choice in the PFC are not time locked to SWRs (whereas the hippocampal representations of upcoming choice are; see Fig 5a and Fig 6a), and not time-locked to hippocampal theta phase (whereas the hippocampal representations are; see Fig 5c and Fig 6c). Finally, the representations of upcoming and alternative choices in the PFC do not show a large overlap in time with the representations in the hippocampus (see updated Fig 4e were we added a statistical test to show the likelihood of the overlap of decoded timepoints). All these results together lead us to hypothesize that SWR-modulation is not the driving factor behind non-local decoding in the PFC.
REVIEWER COMMENT: I do not see how these precisions address my remark. The main claim in the title used to be "Neurons in the medial prefrontal cortex that are not modulated by hippocampal sharp-wave ripples are involved in spatial tuning and signaling upcoming choice." It is now "Neurons in the medial prefrontal cortex are involved in spatial tuning and signaling upcoming choice independently from hippocampal sharp-wave ripples." The substance has not changed. This specific claim is supported solely by Figure 5.
The other analyses cited describe functional characteristics of SWR-unmodulated neurons but, unless linked by explicit new analyses, do not substantiate independence/orthogonality between SWR modulation and non-local decoding in PFC. If there is an analysis that makes this link explicit, it should be clearly presented; as it stands, I cannot find an explanation in the manuscript for why "all these results together" justify the conclusion that "All these results together lead us to hypothesize that SWR-modulation is not the driving factor behind non-local decoding in the PFC". Also: is the main result of this work a "hypothesis"? If so, this should be clearly differentiated from a conclusion supported by results and analyses.
AUTHOR RESPONSE: (3) Based on the reviewers suggestion, we have added a statistical test to compare the phase-locking based of the non-local decoding to hippocampal SWRs and theta phase to shuffled posterior probabilities. Instead of looking at all SWRs in a -2 to 2 second window, we have now only selected the closest SWR in time within that window, and did the statistical comparison in the bin of 0-20 ms from SWR onset. With this new analysis we are looking more directly at the time-locking of the decoded segments to SWR onset (see updated Fig 5a and 6a).
REVIEWER COMMENT: I appreciate the added analysis focusing on the closest SWR and a 0-20 ms bin. My understanding is that you consider the revised analyses in Figures 5a and 6a sufficient to show that predictive non-local representations in mPFC are not linked to hippocampal SWRs and theta phase.
First, the manuscript should explicitly explain the rationale for this analysis and why it is sufficient to support the claim. From the main text it is not possible to understand what was done; the Methods are hard to follow, and the figure legends are not clearly described (e.g. the shuffle is not even defined there).
Specific points I could not reconcile:
i) The gray histograms in the revised Figures 5a and 6a now show a peak at zero lag, whereas in the previous version they were flat, although they are said to plot the same data. What changed?
ii) Why choose a 20 ms bin? A single narrow bin invites false negatives. Please justify this choice.
iii) Comparing to a shuffle is a useful control, but when the p-value is non-significant we only learn that no difference was detected under that shuffle-not that there is no difference or that the processes are independent.
ORIGINAL COMMENT: (2) The main claim of the work is also based on Figure 3, where the authors show that SWRs-unmodulated mPFC neurons have higher spatial tuning, and higher directional selectivity scores, and a higher percentage of these neurons show theta skipping. This is used to support the claim that SWRs-unmodulated cells encode spatial information. However, it must be noted that in this kind of task, it is not possible to disentangle space and specific task variables involving separate cognitive processes from processing spatial information such as decision-making, attention, motor control, etc., which always happen at specific locations of the maze. Therefore, the results shown in Figure 3 may relate to other specific processes rather than encoding of space and it cannot be unequivocally claimed that mPFC neurons "encode spatial information". This limitation is presented by Mashoori et al (2018), an article that appears to be a major inspiration for this work. Can the authors provide a control analysis/experiment that supports their claim? Otherwise, this claim should be tempered. Also, the authors say that Jadhav et al. (2016) showed that mPFC neurons unmodulated by SWRs are less tuned to space. How do they reconcile it with their results?
AUTHOR RESPONSE: The reviewer is right to assert caution when talking about claims such as spatial tuning where other factors may also be involved. Although we agree that there may be some other factors influencing what we are seeing as spatial tuning, it is very important to note that the behavioral task is executed on a symmetrical 4-armed maze, where two of the arms are always used for the start of the trajectory, and the other two arms (North and South) function as the goal (reward) arms. Therefore, if the PFC is encoding cognitive processes such as task phases related to decision-making and reward, we would not be able to differentiate between the two start arms and the two goal arms, as these represent the same task phases. Note also that the North and South arm are illuminated in a pseudo-random order between trials and during cue-based rule learning this is a direct indication of where the reward will be found. Even in this phase of the task, the PFC encodes where the animal will turn on a trial-to-trial basis (meaning the North and South arm are still differentiated correctly on each trial even though the illumination and associated reward are changing).
REVIEWER COMMENT: I appreciate that the departure location was pseudorandomized. However, this control does not rule out that PFC activity reflects motor preparation (left vs right turns) and associated perceptual decision-making/attentional processes that are inherently tied to a specific action. As such, it cannot by itself support the claim that PFC neurons "encode spatial information." Moreover, the authors acknowledge here that "other factors may also be involved," yet this caveat is not reflected in the manuscript. Why?
AUTHOR RESPONSE: Secondly, importantly, the reviewer mentions that we claimed that Jadhav et al. (2016) showed that mPFC neurons unmodulated by SWRs are less tuned to space, but this is incorrect. Jadhav et al. (2016) showed that SWR-unmodulated neurons had lower spatial coverage, meaning that they are more spatially selective (congruent with our results). We have rephrased this in the text to be clearer.
REVIEWER COMMENT: Thanks for clarifying this.
ORIGINAL COMMENT: (3) My reading is that the rest of the paper mainly consists of replications or incremental observations of already known phenomena with some not necessarily surprising new observations:<br /> a) Figure 2 shows that a subset of mPFC neurons is modulated by HPC SWRs and theta (already known), that vmPFC neurons are more strongly modulated by SWRs (not surprising given anatomy), and that theta phase preference is different between vmPFC and dmPFC (not surprising given the fact that theta is a travelling wave).
AUTHOR RESPONSE: The finding that vmPFC neurons are more strongly modulated by SWRs than dmPFC indeed matches what we know from anatomy, but that does not make it a trivial finding. A lot remains unknown about the mPFC subregions and their interactions with the hippocampus, and not every finding will be directly linked to the anatomy. Therefore, in our view this is a significant finding which has not been studied before due to the technical complexity of large-scale recordings along the dorsal-ventral axis of the mPFC.
REVIEWER COMMENT: This finding is indeed non-trivial; however, it seems completely irrelevant to the paper's main claim unless the Authors can argue otherwise.
AUTHOR RESPONSE: Similarly, theta being a traveling wave (which in itself is still under debate), does not mean we should assume that the dorsal and ventral mPFC should follow this signature and be modulated by different phases of the theta cycle. Again, in our view this is not at all trivial, but an important finding which brings us closer to understanding the intricate interactions between the hippocampus and PFC in spatial learning and decision-making.
REVIEWER COMMENT: Yes, but in what way does this support the manuscript's primary claim? This is unclear to me.
ORIGINAL COMMENT: b) Figure 4 shows that non-local representations in mPFC are predictive of the animal's choice. This is mostly an increment to the work of Mashoori et al (2018). My understanding is that in addition to what had already been shown by Mashoori et al here it is shown how the upcoming choice can be predicted. The author may want to emphasize this novel aspect.
AUTHOR RESPONSE: In our view our manuscript focuses on a completely different aspect of learning and memory than the paper the reviewer is referring to (Mashoori et al. 2018). Importantly, the Mashoori et al. paper looked at choice evaluation at reward sites and shows that disappointing reinforcements are associated with reactivations in the ACC of the unselected target. This points to the role of the ACC in error detection and evaluation. Although this is an interesting result, it is in essence unrelated to what we are focusing on here, which is decision making and prediction of upcoming choices. The fact that the turning direction of the animal can be predicted on a trial-to-trial basis, and even precedes the behavioral change over the course of learning, sheds light on the role of the PFC in these important predictive cognitive processes (as opposed to post-choice reflective processes).
REVIEWER COMMENT: Indeed, as I said, the new element here is that the upcoming choice can be predicted. This appears only incremental and could belong to another story; as the manuscript is currently written, it does not support the article's main claim. I would like to specify that, regarding this and the other points above, my inability to see how these minor results support the Authors' claim may reflect my misunderstanding; nevertheless, this suggests that the manuscript should be extensively rewritten and reorganized to make the Authors' meaning clear.
ORIGINAL COMMENT: c) Figure 6 shows that prospective activity in the HPC is linked to SWRs and theta oscillations. This has been described in various forms since at least the works of Johnson and Redish in 2007, Pastalkova et al 2008, and Dragoi and Tonegawa (2011 and 2013), as well as in earlier literature on splitter cells. These foundational papers on this topic are not even cited in the current manuscript.
AUTHOR RESPONSE: We have added these citations to the introduction (line 37).
REVIEWER COMMENT: This is an example of how the Authors fail to acknowledge the underlying problem with how the manuscript is written; the issue has not been addressed except with a cosmetic change like the one described above. The Results section contains a series of findings that are well-known phenomena described previously (see below). Prior results should be acknowledged at the beginning of each relevant paragraph, followed by an explicit statement of what is new, so that readers can distinguish replication from novelty. Here, I pointed specifically to the results of Figure 6, and the Authors deemed it sufficient simply to add the citations I indicated to an existing sentence in the Introduction, while keeping the Results description unchanged. As written, this reads as if these phenomena are being described for the first time. This is incorrect. It is hard to avoid the impression that the Authors did not take this concern seriously; the same issue appears elsewhere in the manuscript, and I fail to see how the Authors "have improved clarity of the text throughout to highlight the novelty of our results better."